![]()
![]()
![]()
T. R. Preston
International Livestock Centre for Africa
P O Box 5689, Addis Ababa, Ethiopia
Introduction
Alternative approaches to the development of feeding systems
Animal response to non-conventional feed resources
Nutritive value
Relating nutrient supply to productive state
Principles of supplementation
Alternative sources of supplements
References
In order to develop feeding systems it is necessary to relate information on the nutritional characteristics of feed resources to the requirements for nutrients according to the purpose and rate of productivity of the animals in question. In the industrialized countries, this information has been incorporated in tables of "feeding standards" which interpret chemical analyses of feed resources in terms of their capacity to supply the energy, amino acids, vitamins and minerals required for the particular productive purpose.
The relevance of feeding standards for developing countries, particularly those in the tropics, has been questioned from the socioeconomic (Jackson 1980) and technical (Graham 1983; Preston 1983) viewpoints. It has been apparent for many years that feeding standards based on assigned nutritive values (e.g. net energy) are misleading when unconventional feed resources are used (e.g. Preston 1972; Leng and Preston 1976; Preston and Leng 1980), since the rates of production achieved may be considerably less than what was predicted. More importantly, this often led to the rejection of many available feed resources which were considered to be too low in digestible energy to support productive functions such as growth and milk production. It also encouraged researchers to copy feeding systems used in temperate countries which are relatively "predictable", but which require feed resources that are unavailable, and/or inappropriate on socio-economic grounds, in most developing countries.
The justification for a new approach to the development of feeding systems for ruminants, not based on conventional "feeding standards", is that:
* The efficiency of the rumen ecosystem cannot be characterised by any form of feed analysis* Feed intake on some diets bears no relationship to digestibility and is much more influenced by supplementation
* Availability of amino acids cannot be inferred from the crude protein content of the diet
* The energy value of a diet, and the efficiency of its utilization, are largely determined by the relative balances of glucogenic energy, long-chain fatty acids and essential amino acids absorbed by the animal
The misconceptions inherent in any system based primarily on feed analysis are that it is almost impossible to predict:
* Whether the feed can support efficient rumen function
* The nature, amounts and the proportions of the end-products of fermentative digestion
* The potential for rumen escape of nutrients and their digestibility in the small intestine.
For these technical reasons, and also because of differing socio-economic circumstances, it has been proposed that a more appropriate objective, especially for developing countries, is to "match livestock production systems with the resources available" (Preston and Leng 1986).
This paper sets out the guidelines for applying these concepts to the development of feeding systems which aim to optimize the utilization of locally available feed resources and to build on traditional practices. Farming systems in developing countries are notoriously difficult to change and innovation must be introduced gradually without inducing excessive risk which may, in the poor conditions of small farmers, directly affect the well-being of the family.
Doubts concerning the usefulness of conventional feeding standards for ruminants in tropical countries surfaced during development work in Cuba (Preston and Willis 1974) in the 1970s when livestock production systems were being established on non-conventional feed resources (i.e. molasses-based diets). In these cases, although nutrient requirements were satisfied, according to traditional feeding standards the responses of the animals did not correspond to the predicted levels of performance.
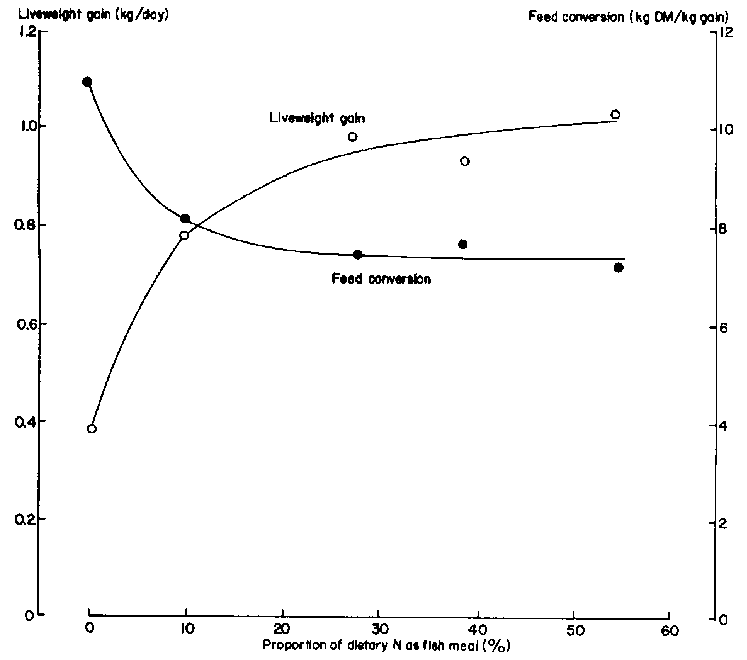
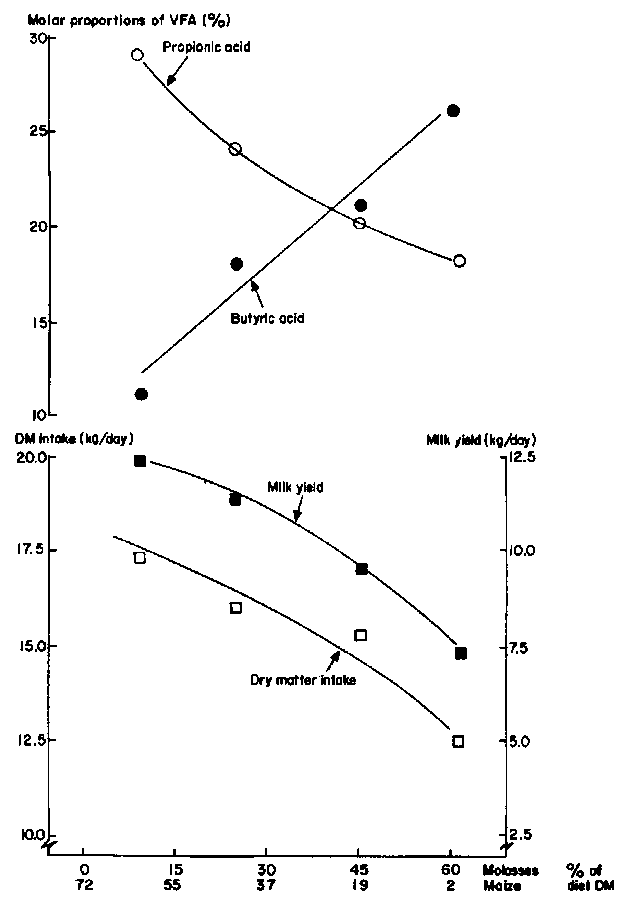
This research demonstrated that small inputs of bypass protein (e.g. Peruvian fishmeal) increased dramatically growth rate and feed efficiency of fattening cattle (Figure 1). In contrast, this feeding system was not able to support high levels of milk production (Figure 2), because of the greater demands of lactation for glucogenic compounds and the relative deficiencies of these in the digestion end-products on molasses-based diets because of the low-propionate, high-butyrate fermentation (Marty and Preston 1970).
The high potential yield of animal products per unit of land devoted to sugar-cane stimulated the subsequent research in Barbados, Mexico, Mauritius and the Dominican Republic aimed at establishing livestock production systems applying the principles which had been developed for feeding molasses (both feed resources have similar concentrations of soluble sugars) (see Preston and Leng 1978a,b).
Research on the feeding value of derinded (Donefer E and James L, cited by Pigden 1972) and chopped (Preston et al 1976) sugar-cane demonstrated that:
* Feed intake was often low even though digestibility was high (6070%)* The animals on this feed apparently needed glucose or glucose precursors because all the sugars are fermented, rumen propionate levels are no higher than observed on high-fibre diets, and the presence of a dense population of ciliate protozoa reduces the availability of microbial protein to the animal (Bird and Leng 1985).
The implication of these two findings is that rumen function did not provide the required balance of nutrients for productive purposes (see Leng and Preston 1976).
Recognition of the role of fermentable N and bypass protein in low-N diets led to research aimed at increasing productivity of cattle and sheep on a range of fibre- and sugar-rich low-N feeds (tong et al 1977; Preston and Leng 1984). Prior to this work, these feed resources were considered to have little value other than to support maintenance and were universally referred to as 'low quality' fibrous feeds.
This led to attempts to improve the digestibility of fibrous feeds by, in particular, alkali treatment (Jackson 1977, 1978). However, the value of alkali treatment was partly obscured by the failure to recognize that the first limitation was not digestibility but the imbalance of nutrients at the level of both the rumen and the whole animal (tong 1982; Preston and Leng 1984).
Combining alkali treatment and appropriate supplementation has led to practical rice-straw-based feeding systems being applied on farms in Bangladesh (Dolberg et al 1981; Davis et al 1983) and Sri Lanka (Perdok et al 1982; Jayasuriya 1984).
The significance of these developments is not so much the use of molasses nor of straw in animal feeding, since both these feeds have been incorporated into diets of ruminants in industrialized countries for many years. The issue is the magnitude of the contribution of molasses and straw to the total dietary dry matter. In industrialized countries their contribution rarely exceeds 10-15% of the diet in the case of molasses, and 20 to 40% for straw, the rest of the ration being cereal grains, highly-fertilized grasses and legumes and oil seed cakes. In contrast, in developing countries the feeding regimes aim to use crop residues and agro-industrial byproducts as the principal component of the diet because these are the locally available resources; and there are restrictions on the use of grain for livestock feeding for financial, political and socioeconomic reasons.
There are many reasons why crop residues, mature pastures and sugar-rich byproducts perform differently when fed as the main component of a diet than when fed as relatively minor components. Some of these differences can be explained on the basis of interactions and associated effects among nutrients, and between nutrients and the site of digestion.
In order that responses in animal productivity to supplements can be predicted accurately on a particular diet, it is necessary to take account of the constraints to metabolism. These are the relative amounts of amino acids, glucogenic energy, VFA energy and long-chain fatty acid energy in the end-products of fermentative and intestinal digestion, since this is what determines the animal's productivity.
Productivity of ruminants is influenced primarily by feed intake which in turn is determined by feed digestibility and the capacity of the diet to supply the correct balance of nutrients required by animals in different productive states. The two major variables that need to be considered are:
* The amounts and balance of nutrients required
* The quantitative availability of nutrients from the diet
The balance of nutrients required depends upon:
* The amounts of dietary components unchanged by rumen fermentation that are absorbed (amino acids, glucose and long-chain fatty acids)* The rates of production of the end-products of fermentative digestion (which can be highly variable)
* Productive functions (pregnancy, lactation, growth, work, maintenance, depletion or repletion of bodyweight)
* Environmental factors (disease, parasitism, temperature and humidity, and other sources of stress)
The availability of nutrients from a diet is highly dependent on:
* The microbial ecosystem in the rumen which influences the availability of microbial protein, VFA energy and glucogenic energy* The chemical composition and physical form of the diet which influences the amounts of protein, starch and long-chain fatty acids which escape the rumen fermentation.
At the present time, it is not possible to match requirements of nutrients for production with the nutrients which become available from digestion, because of the many interactions among the animal, its rumen microbial ecosystem and the diet.
The most widely available low-cost feeds for ruminants in the majority of developing countries are usually native pastures, crop residues and to a lesser extent agro-industrial by- products. Fermentable nitrogen, as urea, is also relatively easy to obtain as it is usually the main source of fertilizer nitrogen. The expensive, and often unavailable (or exported), feeds are the protein meals, derived from oilseed residues and the processing of animals, fish and cereal grains.
The available feed resources are thus primarily sources of potentially fermentable carbohydrates and fermentable nitrogen. This emphasizes the fundamental role played by rumen function in feeding systems in developing countries, especially those in the tropics. The contrast with the industrialized countries is noteworthy since the utilization of their feed resources (dominated by starch-rich cereal grains and protein oilseed meals) depends to a much lesser extent on efficient rumen function. In fact, it is usually advantageous to promote escape of starches and proteins from the rumen so they can be digested in the intestine.
The first step in the strategy to promote efficient utilization of tropical feed resources is to maximize rumen function. This usually means maximizing the rate of degradation of cell wall carbohydrates (the exception is when sugar-rich molasses-based diets are used). Having derived the maximum benefit from rumen fermentation, in terms of the amounts of volatile fatty acids and microbial protein, the next step is to balance these end products of digestion with the amino acids, glucose (and/or its precursors) and long-chain fatty acids which are needed, in varying amounts, for all productive functions such as work, growth, reproduction and lactation.
The potential of the diet to supply these needs will be governed by:
* The pattern of rumen fermentation, since this will determine the relative amounts of microbial protein to VFA that are produced and the proportion of propionic acid which is the main source of glucose;* The amounts of protein, starch and fat in the diet (more usually in the supplement), and the extent to which these nutrients escape the rumen fermentation
The extent to which the protein in a supplement escapes the rumen is partly a function of its rate of degradation (solubility) in the rumen. It is likely to be influenced greatly by the rate of flow of fluid and small particles out of the rumen. This latter characteristic will be influenced by processing of the feed (by physical or chemical means), the presence of some green forage, the amount of protein reaching the duodenum and external factors such as temperature and exercise/work.
The same factors will influence the supply of glucose and glucogenic precursors in terms of the likely bypass of starch to the duodenum. However, the nature of rumen fermentation will have a major influence in terms of the supply of propionic acid.
There is insufficient information available to permit the precise quantification of the proportions of the different nutrients required for different productive states. Nevertheless, an approximation of the needs of animals can be attempted. The suggested scheme attaches relative priorities to the groups of nutrients according to the physiological and biochemical processes underlying the expression of the particular productive state (see Table 1).
The groups of nutrients to be varied for different productive states are:
* VFA energy
* Glucogenic energy
* Amino acids
* Long-chain fatty acids (LCFA)
VFA energy arises from the rumen fermentation of all types of organic matter principally carbohydrates. The principal way of increasing VFA energy in a particular feed is to increase intake and/or the rumen degradability by supplementing with bypass protein and/or alkali treatment (mainly ammoniation).
Table 1. The first limiting nutrients for different physiological functions. These nutrients must be supplied in sufficient quantities and proportions to meet the needs for different functions. When the balance of nutrients is optimum feed intake-is increased and feed utilization efficiency is improved.
|
Physiological function |
Limiting nutrients |
|
Growth (lean tissue) |
Amino acids |
|
Growth (fattening) |
Glucose, LCFA*, amino acids |
|
Puberty |
Glucose, amino acids |
|
Conception |
Glucose, amino acids |
|
Pregnancy |
Glucose, amino acids |
|
Lactation |
Glucose, amino acids, LCFA |
|
Work |
Glucose, LCFA |
* Long-chain fatty acids. Source: Preston and Leng (1986).
Manipulation of the rumen to provide extra protein and glucogenic precursors is still at the experimental stage. Dietary supplementation is the most obvious way of manipulating the supply of absorbed amino acids, glucose and glucose precursors.
Most supplements are expensive and their use in ruminant nutrition competes with monogastric animal and human nutrition. If the primary feed resource is a product of low nutritive value, which would have been wasted if it were not fed to ruminants, it can be argued that the ruminant uses these concentrate supplements more efficiently than monogastric animals. For this reason the term "catalytic" supplement has been used to describe these effects (Preston and Leng 1980). Suckled milk, given in small amounts (<2 litres daily) as a supplement for calves given a straw or molasses-based diet, is a good example of a "catalytic" supplement.
It is mandatory that research should produce response relationships to distinguish economic from biological optima. As a rule of thumb, the role of the supplement ceases to be "catalytic" when it exceeds about 30% of the diet dry matter. Beyond this point it assumes a major role and substitution occurs.
The productive functions of ruminants and the need for supplementary nutrients are discussed in detail by Preston and Leng (1986). The conclusions from their review are outlined below.
Work
Work requires ATP (adenosine triphosphate) generated from the oxidation of long-chain fatty acids, with obligatory requirements for glucogenic compounds and for amino acids (to repair the wear and tear of tissues and replace protein secretions) (see Leng 1985). Since body reserves are a major source of LCFA, it becomes important to allow working animals to fatten in the non-working season.
Maintenance
Maintenance alone obviously requires less energy expenditure than work so there is a proportionately higher demand for amino acids (relative to energy) than in the working animal. This will always be provided by a rumen system which is adequate in fermentable nitrogen.
Growth
Growing animals have a very high requirement for amino acids for tissue synthesis and glucose for oxidation in specific tissues (e.g. brain). In addition considerable amounts of glucose must be oxidized to provide the NADPH required to synthesize fat from acetate. A dietary source of LCFA which can be incorporated directly into fat will spare glucose oxidation.
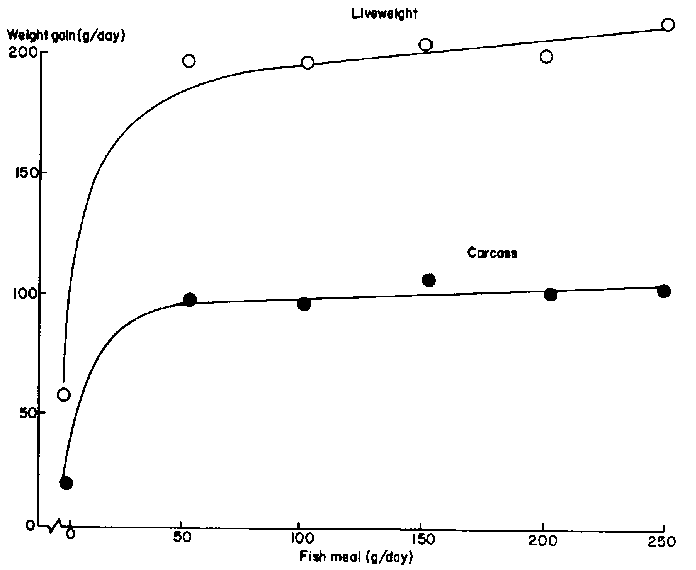
High growth rates cannot be supported on the products of fermentative digestion and bypass protein supplements are essential to take advantage of the VFA energy absorbed. Data taken from Bangladesh and Cuba demonstrate this rationale. Cattle on ammoniated (urea-ensiled) rice straw, when supplemented with only 50 g/d fish meal, increased their liveweight gain threefold (Figure 3). On a molasses-based diet of higher energetic potential, 450 g/d of fishmeal were needed to raise liveweight gain from 300 to 900 g/day (Figure 1).
Reproduction
The supply of glucogenic precursors relative to total energy is an important feature of the improved energy status which results in increased fertility. Recent studies have demonstrated that even when the protein supply is adequate, improvement in the "quality" of the energy (by increasing the glucogenic precursors in the end-products of digestion) accelerated puberty in both males and females (Table 2).
Table 2. Increasing the glucogenic status in the end-products of rumen fermentation (by feeding monensin/lasalocid) of growing bulls and heifers improved their fertility.
|
|
Bulls |
Heifers |
||
|
Control |
Lasalocid |
Control |
Monensin |
|
|
Rumen VFA (% molar) |
|
|
|
|
|
Acetate |
65 |
60 |
74 |
69 |
|
Propionate |
21 |
32 |
19 |
26 |
|
Butyrate |
15 |
7 |
6 |
3 |
|
Testicular volume (cm) |
57 |
91 |
|
|
|
Increase in scrotal circumference (cm) |
3.1 |
5.3 |
|
|
|
Fertility (% cycling) |
|
|
58 |
92 |
Sources: Neuendorff et al (1982); Moseley et al (1982).
Conception rates of cows grazing sub-tropical pasture during the dry season were improved by giving bypass protein (Table 3). A supplement providing fermentable energy (molasses) was much less effective confirming the report of Moseley et al (1982) that it is the "quality" of the energy (i.e. energy in the form of glucogenic compounds) which is the critical issue.
The growth of the conceptus has little effect on the protein and energy demand of ruminants until the last third of gestation when most of the foetal tissues are deposited. It appears that rumen function even on diets of low digestibility can support the birth of a viable offspring of normal weight. This was shown in studies in which urea was included in the drinking water of ewes on nitrogen-deficient pasture (Table 4).
Calf birth weight was increased when pregnant cattle, given a basal diet of hay of low digestibility (45%), were supplemented with urea. However, to prevent bodyweight loss and/or promote weight gain of the dam through pregnancy, it was necessary to provide additional bypass protein (Table 5).
Table 3. Liveweights and conception rates of lactating beef cows grazing native pasture and either a rumen-fermentable carbohydrate (1.86 kg/d of: 85% molasses, 12% cottonseed meal, 17% urea and 1% diammonium phosphate) or a bypass protein meal (1.5 kg/d of cottonseed meal).
|
Supplement |
Liveweight (kg) |
Pregnancy (%) |
|
Nil |
302 |
10 |
|
Rumen-fermentable CHO |
332 |
20 |
|
Bypass protein |
343 |
60 |
Source: Hennessy (1986).
Table 4. Birth weight and growth rate of lambs, and feed and nitrogen intake, wool yield and liveweight loss of ewes grazing low-protein dry pasture.
|
Attribute |
Flinders grass |
Flinders grass + urea (2.2 g urea/litre) |
Flinders grass + urea (1% w/w of grass) |
|
Ewes lambed |
20 |
20 |
20 |
|
Feed intake (g/d) |
900 |
1190 |
1250 |
|
Nitrogen intake (g/d) |
8 |
15 |
18 |
|
Ewe liveweight loss (kg) |
12 |
8 |
9 |
|
Ewes milked |
11 |
- |
10 |
|
Milk yield (ml/4 fur)* |
60 |
ND** |
94 |
|
Lamb survivors |
12 |
16 |
16 |
|
Lamb birth weight (kg) |
2.9 |
3.2 |
3.2 |
|
Lamb growth rate (g/d) |
35 |
81 |
84 |
* Mean yields measured on days 1, 11 and 21
** Not determined Source: Stephenson et al (1981).
It appears that urea supplementation enhances milk production to a level that ensures survival of the offspring by the strategy of giving urea. But to allow the young animal to grow, milk yield must be further stimulated by feeding a bypass protein meal.
Milk production
The major constraint to milk production on diets based on crop residues and agro-industrial byproducts appears to be the availability of glucogenic compounds to provide the glucose for lactose synthesis and for oxidation to provide the NADPH for synthesis of fatty acids.
Table 5. Mean liveweight change and dry-matter intake of pregnant cows (415 kg liveweight) fed spear-grass hay supplemented with urea/sulphur (US) and a bypass protein meal (PP). The experiment was carried out over the last 60 days of pregnancy; the hay was of low digestibility and contained 0.4% N.
|
Diet |
Hay intake (kg DM/d) |
Liveweight change (kg/d) |
Calf birth weight (kg) |
|
Spear grass |
4.2 |
-0.81 |
22 |
|
Spear grass + US* |
6.2 |
-0.31 |
31 |
|
Spear grass + US + PP** |
8.1 |
+0.75 |
32 |
* US supplied 55 g/d N.
** PP supplied 1 kg/d of a protein pellet containing 80% cottonseed meal, 10% fish meal and 10% meat meal (protected with formaldehyde).
Source: Lindsay et al (1982).
There is good evidence that in large ruminants-about 50% of the fatty acids of milk arise from dietary fat. A dietary source of lipid can thus reduce considerably any imbalance caused by relative deficiencies of glucogenic energy and amino acids in the end products of rumen digestion. For many feeding systems in the tropics the level of fat in the diet could be a primary constraint to milk production.
The points to be stressed are that:
* Bypass protein, because of its effects on feed intake, almost always stimulates milk production and depending on the imbalance in nutrients (fermentation pattern) may cause animals to mobilize body reserves. This may be prevented by the use of high-fat, high-protein meals that supply both protein and long-chain fatty acids for digestion post ruminally.* Bypass starch or manipulation of the rumen to give higher propionate production (e.g. by supplementation with monensin), because it balances nutrients for milk production, may prevent mobilization of body reserves without large effects on feed intake and therefore on milk production. But because it balances the nutrients for milk production, efficiency of energy utilization is increased and bodyweight is often increased.
Wool or hair production
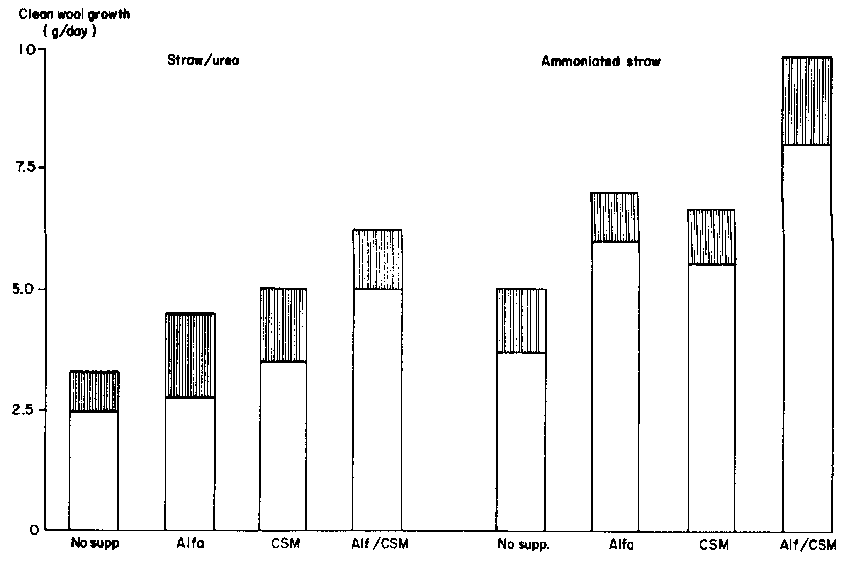
The effect of nutrition on wool production appears to be dependent almost entirely on the quantity, and balance, of the amino acids absorbed. Therefore, feed intake is the primary limitation to wool or fibre growth. Improving the balance of protein relative to energy in the products of digestion (e.g. removing protozoa from the rumen, supplying a rumen "activator" and/or giving bypass protein) will increase wool and fibre growth (Figure 4).
The proposed scheme (Table 6) is empirical but is considered to be appropriate for the conditions of most developing countries.
Select the basal carbohydrate-rich resource
The first step is to select the basal carbohydrate resource according to availability, potential fermentability and price. Supplementary nutrients should then be provided in accordance with their relative priorities and costs.
Fermentable N
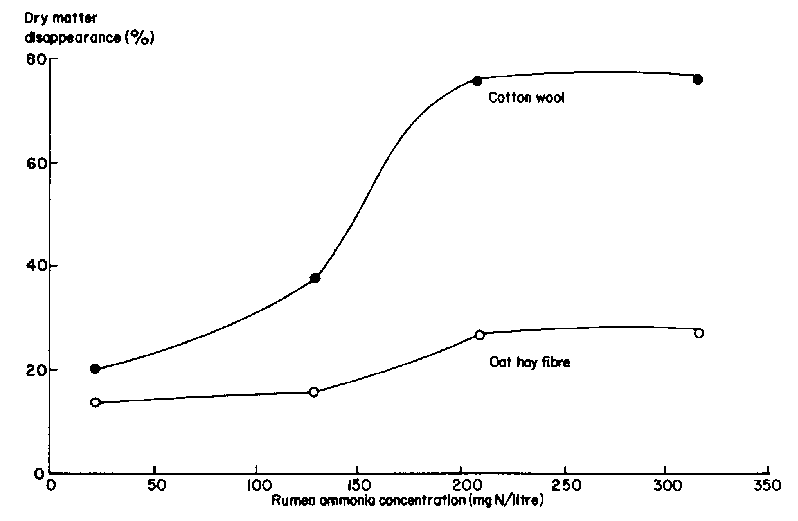
The first supplement to be considered should be a source of fermentable nitrogen (usually urea or ammonia) to ensure the level of rumen ammonia is above 150 mg/litre of rumen fluid. The generally recommended minimum level of rumen ammonia to support efficient use of fermentable carbohydrate for microbial growth is 50 mg/litre. However, this appears to be too low to optimize the rate of degradation of fibrous substrate, since the disappearance rate of cellulose and fibre from nylon bags in the rumen was increased when the concentration of ammonia was raised to 200 mg/litre (see Figure 5).
Table 6. Priorities for nutritional supplements to optimize the utilization of diets based on crop residues, sugar-rich agro-industrial byproducts or high-biomass producing tropical crops.
1: Select the carbohydrate-rich resource which will be the basis of the diet.
2: Ensure the rumen ecosystem is optimum by:
(i) Providing fermentable N (a total of about 3 g N/100 g fermentable carbohydrate) to raise rumen amonia levels to at least 15 mg/100 ml rumen fluid.(ii) Supplying a highly digestible (preferably green) forage
3: Balance the end-products of digestion by:
(i) Manipulating the rumen (increase propionate, accelerate flow rate out of the rumen, reduce or eliminate protozoa)(ii) Supplementing with sources of bypass nutrients to improve protein: energy ratio and raise the glucogenic status of the diet in accordance with animal needs:
* Protein
* Starch
* Long-chain fatty acids (LCFA)
Source: Preston and Leng (1986).
When the substrate was alkali-treated maize cobs (Alvarez et al 1983), rate of dry matter loss from nylon bags in the rumen increased linearly as rumen ammonia concentration was raised from 30 to 120 mg/litre of rumen fluid.
Rate of degradation of cell wall carbohydrate is of paramount importance when the diet is based on crop residues; because it is the rate of degradation of fibre which eventually limits feed intake and therefore animal productivity.
When rumen ammonia levels are lower than 150 mg/litre it is recommended that the effects of adding urea should be monitored under the prevailing field/farm situation. As a general rule, if a deficiency is suspected, urea should be added at the rate of about 1-2% of the organic matter in the diet. It is desirable that supplementation ensures an almost continous supply of ammonia-nitrogen in the rumen and the use of molasses/urea blocks or high-urea (10%) liquid mixtures with molasses is a convenient way of ensuring this. Ammoniation using ammonia gas, or through ensiling with urea, are other ways of proving a continuous supply of rumen ammonia with the associated advantage of upgrading the carbohydrate component. A recent development is the generation of ammonia from mixtures of dry chemicals (e.g. ammoniate sulphate and quicklime) when these are mixed with water (Mason et al 1985).
Highly digestible forage
The second supplement should be a source of highly digestible forage, preferably legume (or a byproduct such as beet pulp) given at about 10-20% of the diet. The exact action of this type of supplement on rumen function is not fully understood. In some way it helps to ensure a more efficient rumen environment for the digestion of cell wall carbohydrate (Table 7) perhaps by providing micro-nutrients (e.g. peptides, amino acids, minerals, vitamins) which increase fungal biomass and/or the rate of bacterial colonization of the fibre.
Table 7. Dry matter disappearance from nylon bags of straw incubated in the rumens of sheep fed on untreated straw or straw supplemented with sugar beet pulp.
|
|
Diet of host sheep |
||
|
Untreated straw |
Straw plus 150g beet pulp/kg stew |
||
|
Rumen pH |
7.0 |
6.9 |
|
|
Rumen NH3 (mg/litre) |
151 |
129 |
|
|
Dry matter loss (%) |
|
|
|
|
|
In 24 hr |
39 |
41 |
|
|
In 48 hr |
46 |
50 |
|
|
In 72 hr |
51 |
55 |
Source: Silva and Ørskow (1985).
Bypass protein
The third supplement should be an oilseed meal, cereal bran or an animal byproduct meal (supplying protein and fat) and should be given in amounts not to exceed 30% of the total diet dry matter. The 30% limit is to prevent depression/substitution of the digestible energy of the basal diet. Lesser amounts may be more economical, and it is imperative that feeding trials be carried out to define response relationships. In this way the amount of supplement can be related to the rate of animal productivity. The optimum level (in economic rather than biological terms) and the degree of response to the supplement, will depend upon the fermentability of the basal diet.
Long-chain fatty acids (LCFA)?
Supplementation with a source of long-chain fatty acids (LCFA) is a strategy that promises to be of considerable benefit, especially on diets with a low content of lipids (e.g. crop residues and molasses). However, more research is needed before making recommendations.
The limiting nutrients for production on most tropical feed resources are fermentable nitrogen, glucogenic precursors and bypass protein and dietary long-chain fatty acids. Urea, oilseed cakes, cereal milling byproducts and animal byproduct meals are the logical supplements when available. However, there are many situations where farmers do not have access to these supplements either because they are not locally available or are too expensive. In addition, there is often a reluctance to use urea because of the fear of toxicity.
Livestock excreta
Excreta from all types of livestock have been used in livestock rations. It is obvious that excreta in general must be a poor source of fermentable carbohydrate and protein. However, long-chain fatty acids may build up in litter as small amounts will be present in faeces and they are only slowly degraded by microorganisms in the litter. Microbial growth in the litter will therefore tend to concentrate these fatty acids.
Excreta from ruminant animals are high in refractory cell-wall carbohydrate with smaller amounts of microbial cells (from the caecum) and some urea if the urine is incorporated with the faeces. The monogastric species produce the most valuable excreta; and especially in the case of poultry, there may be considerable contamination with wasted feed grains. Excreta from poultry is rich in nitrogen mostly as uric acid which is hydrolyzed to ammonia by rumen micro-organisms.
Excreta (often depathogenized with formalin or by ensiling) have been used widely in the developed countries as a component of cereal-grain-based diets in which their main contribution is as a source of non-protein nitrogen and minerals.
In developing countries, only poultry litter has found ready acceptance as a component of livestock feeds. It appears to play a particularly appropriate role in high-molasses diets, where it complements the readily fermentable sugars and the low levels of fermentable N and of phosphorus. There appears to be a particularly beneficial effect of poultry litter on rumen propionate production in cattle fed a molasses-based diet (Fernandez and Hughes-Jones 1981; Marrufo 1984). It is well documented that this is reflected in higher levels of animal performance (Meyreles and Preston 1982; Meyreles et al 1982).
The data in Table 8 show that poultry litter is less effective than fishmeal or an oil cake meal for supplementing cattle given a molasses- or a pasture-based diet, from which it can be inferred that it provides little or no bypass protein. This is to be expected in view of its chemical characteristics.
Table 8. Poultry litter is a less effective supplement than fish meal (in a molasses diet) or sunflower cake (on pasture), indicating that it supplies little or no bypass protein.
|
On pangola pasture (plus 1.5 kg/d of molasses-urea): | ||||
|
|
Amount of supplement (kg/d) | |||
|
Poultry litter |
1.3 |
1.0 |
0.64 |
0.0 |
|
Sunflower cake |
0.0 |
0.12 |
0.31 |
0.6 |
|
Liveweight gain (g/d) |
480 |
580 |
680 |
740 |
|
Diet of molasses-urea ad libitum and restricted forage: |
||
|
|
N-supplement |
|
|
Poultry litter |
Fish meal |
|
|
Liveweight gain (g/d) |
575 |
825 |
Sources: Delgado et al (1977); Preston et al (1970).
Legume forages and foliages from food crops
An alternative resource which can serve as a source of fermentable N and of bypass protein is a forage crop grown on the farm, or produced as a byproduct or residue from a food crop. A legume crop has a further advantage because of its capacity to fix atmospheric nitrogen and thus spare the need for fertilizer N. Based on the premise of using protein economically, then the intake of these forages should be restricted and therefore it is preferable to grow the legume as a pure sward.
The amount of bypass protein in a legume (green or dry and used as a supplement) has not been estimated. It may be beneficial to grow tannin-rich legumes when these are to be used as supplements to crop residues.
The tree legumes such as gliricidia, erythrina and leucaena have very great potential because they are high yielding and perennial. They are also deep-rooted and may have access to water and nutrients (e.g. phosphorus) unavailable to smaller plants. In the tropics, tree legumes have a special role, since they can also be used for shade (e.g. in coffee plantations), as "live" fences and as sources of fuel. Some of them (e.g. glyricidia and erythrina) can be established easily from cuttings hence their use as "live" posts.
The other valuable forages are from cassava and sweet potatoes and, to a lesser extent, bananas. Some results from using these materials as supplements in molasses-based diets are described by Ffoulkes and Preston (1978).
Attributes of legumes as supplements
In developing countries where competition for land to grow crops or grazing is high, the area likely to be sown to fodder legumes will be almost always a small proportion of the total. It follows therefore that the role of a legume must be to increase the efficiency of utilization of the basal diet (i.e. a low-N pasture or a crop residue) at low levels of supplementation (usually less than 20%) and used "catalytically".
As a priority, the legume should have a high protein content to supply both fermentable and bypass protein; there will be additional benefits if the legume contains other critical nutrients (e.g. lipids, minerals, vitamins and other plant compounds) which enhance the rumen ecosystem so as to increase microbial growth, rate of fibre digestion, propionate production and escape of dietary protein.
It is likely that legume forages rich in tannins will be superior as sources of bypass protein since tannins link with proteins during mastication, and appear to reduce microbial degradation of plant proteins (Reid et al 1974). The high levels of tannins in Lotus pedunculatus, whilst protecting protein from degradation, reduce digestibility of fibre by inhibiting the activity of bacteria (Chesson et al 1982) and fungi (Akin and Rigsby 1985). Barry (1985) considered that the ideal concentration of condensed tannins was 20-40 g/kg diet dry matter; and that higher levels (76-90 g/kg) were detrimental. He also found that sheep could adapt to high tannin levels. Provided that tannin-rich plants are only used as supplements (e.g. less than 25% of the diet dry matter), there is unlikely to be a serious problem and their presence in the diet may well be beneficial (Barry and Manley 1984). Examples of tropical legumes which are known to contain tannins are: leucaena, glyricidia and sesbania.
In situations where the fermentable N requirement can be met from other sources (e.g. urea or animal excreta) the need is to reduce the degradability of the legume protein so as to increase its bypass characteristics. This has been shown to occur when a forage is artificially dried and more so when pelleted (see Table 9). Rarely will it be economic to dehydrate or pellet legume forages and sun-drying is the only feasible alternative.
Table 9. The bypass properties of clovers are increased when the herbage is frozen or dried and pelleted.
|
|
Fresh |
Frozen |
Artificially dried/pelleted |
|
Amino acid intake (g/d) |
127 |
127 |
124 |
|
Amino acids entering small intestine (g/d) |
80 |
133 |
175 |
Source: Preston and Leng (1986).
As discussed earlier, secondary plant compounds such as tannins are known to protect dietary proteins against rumen microbial attack. Thus if a freshly harvested or grazed legume is to be used as a bypass protein supplement then it should be selected for a relatively high content of tannins.
This point is illustrated by the data in Table 10 which show that although the tannin-containing legumes (trefoil and sanfoin) were less palatable than lucern, nevertheless they supported faster growth rates in heifers. The authors concluded that this was because more of the protein in the legumes containing tannins escaped degradation in the rumen.
Table 10. Relative palatability of three legumes, and heifer growth performance when grazing them.
|
|
Alfalfa |
Trefoil |
Sainfoin |
|
Relative palatability* |
34 |
46 |
45 |
|
Liveweight gain (g/d) |
670 |
810 |
800 |
* Percentage rejection of forage on offer.
Source: Marten and Ehle (1984).
Tropical legumes generally are richer in tannins than are temperate legumes and therefore should function better as sources of bypass protein. Evidence for this is provided by the results of feeding trials with tropical tree legumes discussed earlier. It must be emphasized that when legumes contain a high proportion of protected protein, then some other source of rumen fermentable nitrogen will be required, usually urea.
Akin D E and Rigsby L L. 1985. Influence of phenolic acids on rumen fungi. Agronomy Journal 77:180-182.
Alvarez F. Dixon R M and Preston T R. 1983. Ammonia requirements for rumen fermentation. In: Farrel D J and Pran Vohra (eds). Recent advances in ruminant nutrition in Australia 1983. University of New England Publishing Unit, Armidale, New England, Australia. p. 9A.
Barry T N. 1985. The role of condensed tannins in the nutritional value of Lotus pedunculatus for sheep 3: Rates of body and wool growth. British Journal of Nutrition 54:211-217.
Barry T N and Manley T R. 1984. The role of condensed tannins in the nutritional value of Lotus pedunculatus for sheep 2: Quantitative digestion of carbohydrates and proteins. British Journal of Nutrition 51:493-504.
Bird S H and Leng R A. 1985. Productivity responses to eliminating protozoa from the rumen of sheep. In: Leng R A, Barker J S F. Adams D and Hutchinson K (eds). Biotechnology and recombinant DNA technology in the animal production industries in Australia. Reviews in Rural Science No. 6. University of New England Printing Unit, Armidale, New England, Australia.
Chesson A, Stewart C S and Wallace R J. 1982. Influence of plant phenolic acids on growth and cellulolytic activity of rumen bacteria. Applied Environmental Microbiology 44:597-603.
Davis C H. Saadullah M, Dolberg F and Haque M. 1983. Ammonia treatment of straw for cattle production in intensive agrarian agriculture. In: Davis C H. Preston T R. Haque M and Saadullah M (eds). Maximum livestock production from minimum land. Bangladesh Agricultural University, Mymensingh. pp. 1-25.
Delgado A, Elias A, Veitia J L and Garcia R. 1979. The use of pasture for beef production. 6. Poultry litter: runflower cake meal ratios in the supplementation of bulls grazing pangola grass during the dry season. Cuban Journal of Agricultural Science 13:265-272.
Dolberg F. Saadullah M and Haque M. 1981. Straw treatment in a village in Noakhali district, Bangladesh. In: Jackson M G. Dolberg F. Davis C H. Haque M and Saadullah M (eds). Maximum livestock production from minimum land. DANIDA, Copenhagen. pp. 205-224.
Fernandez A and Hughes-Jones M. 1981. Rumen fermentation and rumen function in bulls receiving a basic diet of molasses/urea supplemented with poultry litter, sweet potato forage or wheat bran. Tropical Animal Production 6:360.
Ffoulkes D and Preston T R. 1978. Cassava or sweet potato forage as combined sources of protein and roughage in molasses-based diets: Effect of supplementation with soybean meal. Tropical Animal Production 3:186-192.
Graham N McC. 1983. Feeding standards: An outmoded concept in ruminant nutrition. In: Farrel D J and Pran Vohra (eds). Recent advances in ruminant nutrition in Australia 1983. University of New England Publishing Unit, Armidale, New England, Australia. pp. 184-191.
Hennessy D W. 1986. Supplementation to ameliorate lactational anoestrous in first-calf heifers grazing native pastures in the subtropics. Animal Production in Australia 17 (submitted).
Jackson M G. 1977. The alkali treatment of straws. Animal Feed Science and Technology 2:105-130.
Jackson M G. 1978. FAO Animal Production and Health Paper No. 10. FAO, Rome.
Jackson M G. 1980. Who needs feeding standards? Animal Feed Science and Technology 6:101-104.
Jayasuriya M C N. 1984. Potential for the better utilization of crop residues and agro-industrial by-products in animal feeding in the Indian sub-continent. In: Preston T R. Vappu V Kossila, Goodwin J and Sheila Reed (eds). Guidelines for research on the better utilization of crop residues and agro-industrial by-products in animal feeding in developing countries. 1. State of knowledge. FAO, Rome.
Leng R A. 1982. A theoretical discussion on the factors limiting production in cattle fed basal diets of straw. In: Preston T R. Davis C H. Dolberg F. Haque M and Saadullah M (eds). Maximum livestock production from minimum land. Bangladesh Agricultural University, Mymensingh. pp. 79-104.
Leng R A. 1985. Muscle metabolism and nutrition in working animals. In: Proceedings of the ACIAR workshop on draught animal power for production. James Cook University, Townsville, Australia. (In press).
Leng R A and Preston T R. 1976. Sugar cane for cattle production: Present constraints, perspectives and research priorities. Tropical Animal Production 1:1-22.
Leng R A, Kempton T J and Nolan J V. 1977. Nonprotein nitrogen and bypass proteins in ruminant diets. AMRC Review 33:1-21.
Lindsay J A, Mason G W J and Toleman M A. 1982. Supplementation of pregnant cows with protected proteins when fed tropical forage diets. Proceedings of the Australian Society of Animal Production 14:67-78.
Marrufo D. 1984. La Leucaena leucocephala: su productividad en la zona henequenera de Yucatan y su uso como suplemento en dietas a base de melaza/urea. Tesis de Maestria, Universidad de Yucatan.
Marten G C and Ehle F R. 1984. Influence of quality variation in four legume species on weight gains of grazing heifers. Agronomy Abstracts p. 159.
Marty R J and Preston T R. 1970. Molar proportions of the short chain volatile fatty acids (VFA) produced in the rumen of cattle given high-molasses diets. Revista Cubana Ciencia Agricola 4:183-187.
Mason V C, Cook J E, Smith T. Siviter J W. Keene A S and Hartley R D. 1985. The new AGRI-AM process for ammoniation: Stack ammoniation of mature forages using ammonium salts. In: Preston T R. Nuwanyakpa M, Mason V C and Lambourne L J (eds). Proceedings of the 2nd annual workshop of the African Network for Agricultural Byproducts. ILCA, Addis Ababa, Ethiopia.
Meyreles L and Preston T R. 1982. The role of poultry litter in molasses/urea diets for fattening cattle. Tropical Animal Production 7:138-141.
Meyreles L, Pound B and Preston T R. 1982. The use of Leucaena leucocephala or sugar cane tops as sources of forage in cattle diets based on molasses/urea, supplemented with chicken litter and/or wheat bran. Tropical Animal Production 7:92-97.
Moseley W M, McCartor M M and Randel R D. 1982. Effects of monensin on growth and reproductive performance of beef heifers. In: Randel R D (ed.). Nutritional influences on reproductive development of replacement heifers. Technical Report 82:1-21.
Neuendorff D A, Rutter L M, Peterson L A and Randel R D. 1982. Effect of lasolacid on growth and puberal development. In: Randel R D (ed.). Nutritional influences on reproductive development of replacement heifers. Technical Report No. 1, Texas A & M University.
Perdok A B. Thamotaaram M, Blum J J. van den Born H and van Velun C. 1982. Practical experiences with urea-ensiled straw in Sri Lanka. In: Preston T R. Davis C H. Dolberg F. Haque M and Saadullah M (eds). Maximum livestock production from minimum land. Bangladesh Agricultural University, Mymensingh.
Pigden W J. 1972. Sugar cane as livestock feed. Report to the Carribean Development Bank, Barbados.
Preston T R. 1972. Molasses as an energy source for cattle. World Review of Nutrition and Dietetics 17:280-311.
Preston T R. 1983. Feeding standards can be misleading. In: Farrell D J and Pran Vohra (eds). Recent advances in animal nutrition in Australia. University of New England Publishing Unit, Armidale, Australia.
Preston T R and Leng R A. 1978a. Sugar cane as cattle feed. I: Nutritional constraints and perspectives. World Animal Review 27:7-12.
Preston T R and Leng R A. 1978b. Sugar cane as cattle feed. II: Commercial application and economics. World Animal Review 28:448.
Preston T R and Leng R A. 1980. Utilization of tropical feeds by ruminants. In: Ruchebush Y and Thivend P (eds). Digestive physiology and metabolism in ruminants. MTP Press, Lancaster. pp. 621-640.
Preston T R and Leng R A. 1984. Supplementation of diets based on fibrous residues and byproducts. In: Sundstol F and Owen E (eds). Straw and other fibrous byproducts as feed. Elsevier Press, Amsterdam, the Netherlands. pp. 373-413.
Preston T R and Leng R A. 1986. Matching livestock production systems to available resources. ILCA, Addis Ababa, Ethiopia. 331 pp.
Preston T R and Willis M B. 1974. Intensive beef production. Second edition. Pergamon Press, Oxford, UK.
Preston T R. Carcano C, Alvarez F J and Guttierez D G. 1976. Rice polishings as a supplement in a sugar cane diet: Effect of level of rice polishings and processing the sugar cane by derinding or chopping. Tropical Animal Production 1:150-163.
Reid C W S. Ulyatt M J and Wilson J H. 1974. Plant tannins, bloat and nutritive value. Proceedings of the New Zealand Society of Animal Production 34:82-92.
Silva A and Ørskov E R. 1985. Effect of unmolassed sugar beet pulp on the rate of straw degradation in the rumens of sheep given barley straw. Proceedings of the Nutrition Society 44:50A.
Stephenson R C A, Edwards J C and Hopkins P S. 1981. The use of urea to improve milk yield and lamb survival of merinos in a dry tropical environment. Australian Journal of Agricultural Research 32:497-509.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}