![]()
![]()
![]()
Abstract
Introduction
Materials and methods
Results and discussion
Conclusions
Acknowledgements
References
R.J. Chamberlin1, the Late D.C.I. Peake1, R.L. McCown2, I. Vallis3, and R.K. Jones4
CSIRO Australia, Division of Tropical Crops and Pastures
1 CSIRO Laboratories, Darwin, Northern Territory 5789.
2 CSIRO Laboratory, Townsville, Queensland 4814.
3 CSIRO Cunningham Laboratory, St. Lucia, Queensland 4067.
4 ACIAR/CSIRO Project, c/o Australian High Commission, Nairobi, Kenya.
Growth and yield of maize as a sole crop was compared with that of maize with intercrops of Alysicarpus vaginalis, stylosanthes hamata cv. Verano, or Centrosema pascuorum under conditions in which soil water was adequate prior to physiological maturity of the crop. Data from three seasons are presented. In the first season, when only one N rate was used with a range of legume densities, maize yield varied inversely with legume yield. In the other two seasons N rate was varied, and 15N was used to obtain more information on the competitive relationships between the components of the intercropping systems. Results show that when conditions are good for establishment and early growth of the legumes, competition between the legume intercrop and maize occurs. This effect was greatest at low N rates and less with Verano stylo than with A. vaginalis. There was never any evidence of a positive contribution by any of the legumes to the N economy of the maize.
Legume seed production, essential for regeneration of the legume in the cropping system being studied, varied with species and seasonal conditions.
This research is part of a project on the feasibility of a no-tillage, ley farming system for the semi-arid tropics of northern Australia (McCown et al 1985). When a cereal crop is sown after a legume phase in such a system one can either prevent establishment of the legume from seed using pre-emergence herbicides or allow the legume to grow as an understory in the cereal crop. Advantages of the latter approach include (a) improvement of the quality of residues for cropland grazing during the dry season, (b) provision of additional seed for subsequent legume pasture establishment, and (c) protection against soil erosion by the legume cover.
Legumes in close association with nitrophilous crops have increased crop production (Nair et al 1979; IITA 1980; Waghmare and Singh, 1984), but other studies have shown that legumes can compete for soil N with the associated crop and reduce its grain yield (Kurtz et al 1952a,b; Enyi, 1973; Wahua, 1983). Our research aims to elucidate the interactions between a maize crop and a forage legume understory under (a) nitrogen-limiting conditions and (b) under water-limiting conditions, so that the requirements for maintenance of high intercrop maize yields can be defined. This paper reports findings on the N aspects when water was non-limiting.
All experiments were carried out at CSIRO's Katherine Research Station, northern Australia, latitude 14° S. longitude 132° E. The soil is a red earth (Oxic Paleustalf) (Williams et al 1985). Fertilizer was applied so that all elements except N were non-limiting. Three well-adapted legumes differing in growth habit and maturity were used as intercrops. All behave as annuals in this climate. Stylosanthes hamata cv Verano is semi-erect and is intermediate in maturity; Alysicarpus vaginalis (Martin and Torssell 1974) is semi-erect and the earliest maturing; Centrosema pascuorum has twining stems and is late maturing (Clements et al, 1983).
In all experiments maize was sown at a row spacing of 75 cm. Mid-season cultivars that reached anthesis approximately 50 days after sowing and reached maturity after approximately 100 days were used. Legumes were sown at the same time as the maize, and all plots were hand weeded.
Experiment 1 was designed to test whether a legume understory reduced maize grain yield. In the 1979/80 wet season the maize cultivar DeKalb XL99 Was sown to give a population of 45,000 plants/ha. Legumes were broadcast at rates shown in Table 1. The design was a split plot with legume species as main plots and seeding rates as subplots. Subplot size was 3 m x 5 m and there were two replicates. Ammonium nitrate fertilizer was applied at 75 kg N/ha broadcast 2 weeks before sowing and 30 kg N/ha broadcast at sowing. The crops were not irrigated and the rate of N applied was expected to give maize grain yields of 5-6 t/ha. At physiological maturity maize grain yield was measured from an area of 4.5 m2 and legume dry matter yield from two 0.25 m2 areas. A further 0.5 m2 of legume was sampled at legume maturity when seed yield was also measured.
Table 1. Legume seeding rates, Experiment 1.
|
Species |
Seeding rate (kg/ha) |
|||
|
Density 1 |
Density 2 |
Density 3 |
Density 4 |
|
|
S. hamata |
1 |
8 |
30 |
180 |
|
A. vaginalis |
1 |
6 |
20 |
80 |
|
C. pascuorum |
2 |
10 |
35 |
120 |
Experiment 2 was carried out in the 1982/83 wet season and compared maize (cv. Sergeant) grown as a sole crop with maize intercropped with A. vaginalis, S. hamata, or C. pascuorum. Maize population was 60,000 plants/ha and to simulate a natural situation legume seed was broadcast on appropriate plots at rates sufficient to give stands of approximately 1.5 million plants/ha. Water deficits were prevented by a supplementary irrigation schedule based on pan evaporation measurements. Three rates of N were applied as ammonium sulphate: low - 25 kg N/ha broadcast at sowing; medium (marginally adequate) - 50 kg N/ha broadcast at sowing; and high - 50 kg N/ha broadcast at sowing, 40 kg N/ha banded 25 days after sowing and 50 kg N/ha broadcast at silking. Rain after the first broadcast application washed the fertilizer into the soil. The experiment was a split plot design with N rates as main plots, and legume species as subplots. There were two replicates with sub plot size of 6 m x 6 m. Ammonium sulphate enriched with 15N was applied in solution to microplots within the low and medium N treatments at sowing at a rate of 50 mg 15N/m2. Microplot size was 2.0 m x 1.25 m with a sample area of 1.5 m x 0.75 m. At maize maturity total legume yield, total maize yield (including senescent leaves), maize grain yield and components of yield were measured. Sample size for maize was 11.25 m2, and 2.25 m2 for the legume. Plants from the microplots were also harvested at maize maturity. All samples were analysed for total N. and those from the microplots were also analysed for 15N.
Experiment 3 was carried out in the 1983/84 wet season. Maize cultivar and cultural and sampling methods were the same as in Experiment 2, but the climbing legume C. pascuorum was not included. Nitrogen rates were: 25 kg/N ha broadcast at sowing (N1); 50 kg N/ha broadcast at sowing (N2); 50 kg N/ha broadcast at sowing and 50 kg N broadcast 30 days after sowing (N3); 100 kg N/ha broadcast at sowing and 100 kg N/ha broadcast 30 days after sowing (N4). The fertilizer was watered into the soil by rain after the first application and by irrigation after later applications. The experiment was a split plot design with N rates as main plots and legume species as subplots. There were 4 replicates and subplot size was 14 m x 3 m. As in Experiment 2 N was applied as ammonium sulphate and irrigation maintained an adequate water supply. Fertilizer enriched with 15N was applied to microplots in the N1, N2, and N3 treatments which were sampled at 30 days after sowing, at silking and at maize maturity. The rate of 15N application and microplot size were the same as in Experiment 2. Yield measurements at maize maturity were from 5 m2 for maize and 2.25 m2 for the legumes. Chemical analyses were similar to those of Experiment 2. The proportion of photosynthetically active radiation reaching the legume understory was measured with a Licor line sensor on two occasions during maize grain filling (10 and 24 days after Bilking). The mean value of radiation received at five positions at the top of the legume canopy in each plot was expressed as a proportion of that above the maize crop.
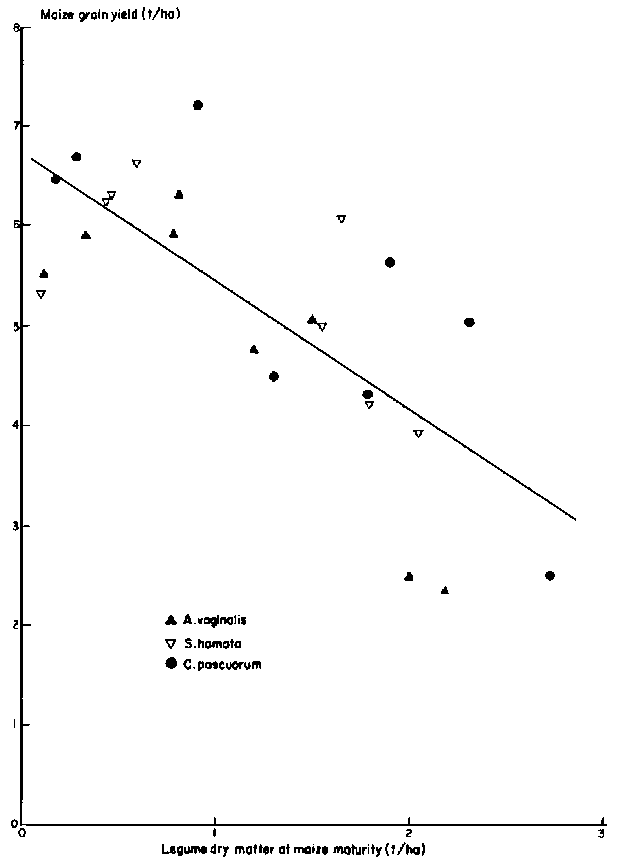
Under the cultural and environmental conditions of Experiment 1, maize grain yield decreased as legume production increased (Figure 1). There were no significant differences among species and the common regression through all points accounted for 56 % of the variance. The reduction in grain yield (1.28 t of grain forgone for each tonne of forage produced) is economically unacceptable.
Meteorological records for this season show that rainfall was above average and well distributed, and although soil water was not monitored, water balance simulations confirm that the water supply to the plants in this season was very favourable. CORNF (Stapper and Arkin, 1980) was used for this calculation, and the effect on water use of increased leaf area due to intercropping was approximated by increasing maize plant density and leaf area per plant in the model. Simulated available soil water was high throughout the season and declined to 30 - 35 mm at maize maturity. It seems unlikely that water shortage accounted for the depression of maize yield by intercrops.
Figure 1. The effect of yields of intercropped legumes on Brain yield of maize, Experiment 1.
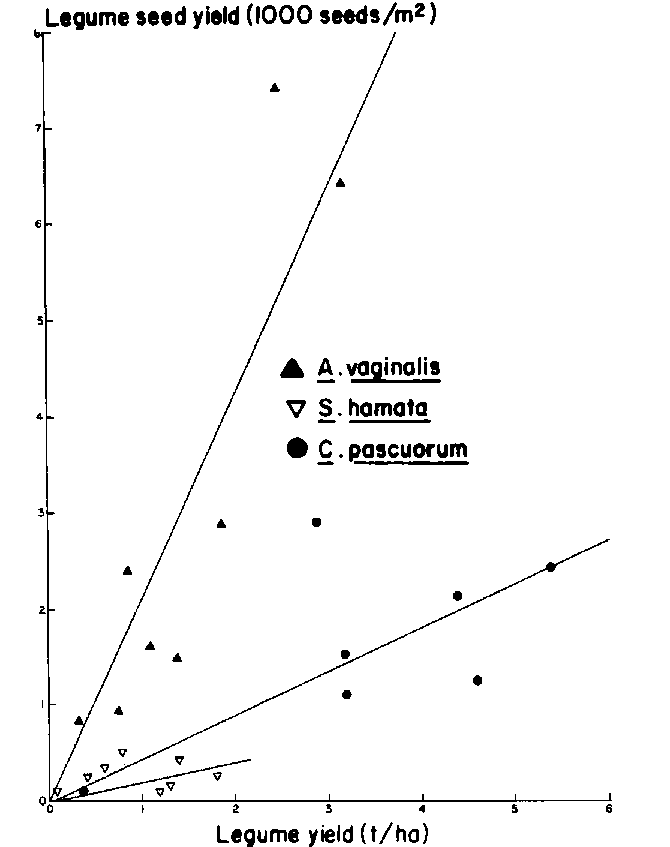
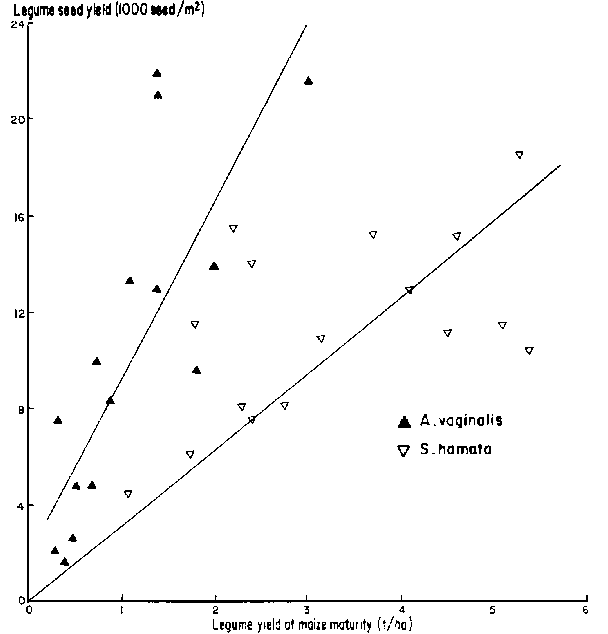
Seed yield of all legumes was low (Figure 2) but, except for S. hamata, was sufficient for re-establishment. Argel and Humphreys (1983) have shown flowering and inflorescence development of S. hamata to be sensitive to decreased irradiance.
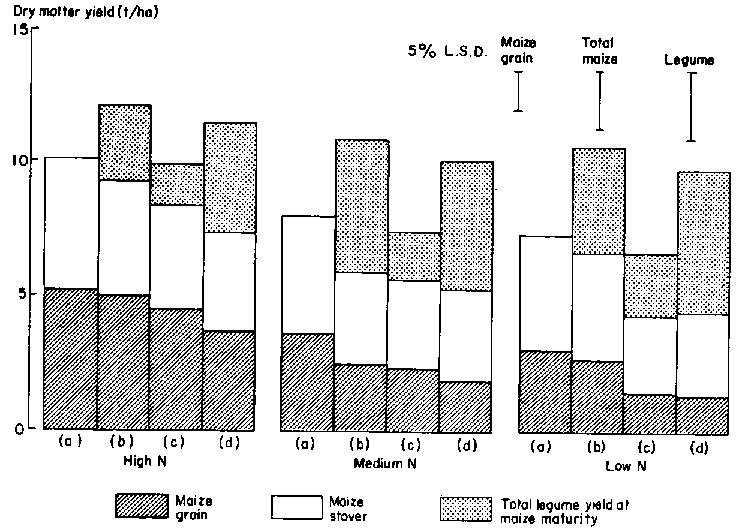
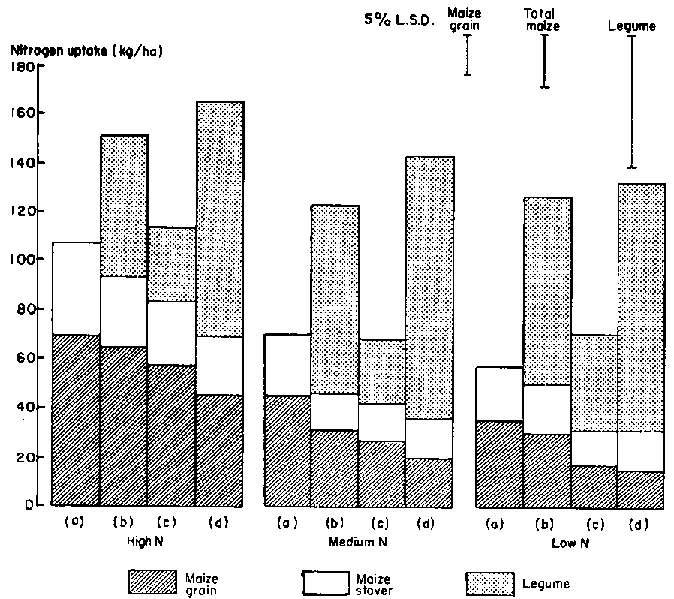
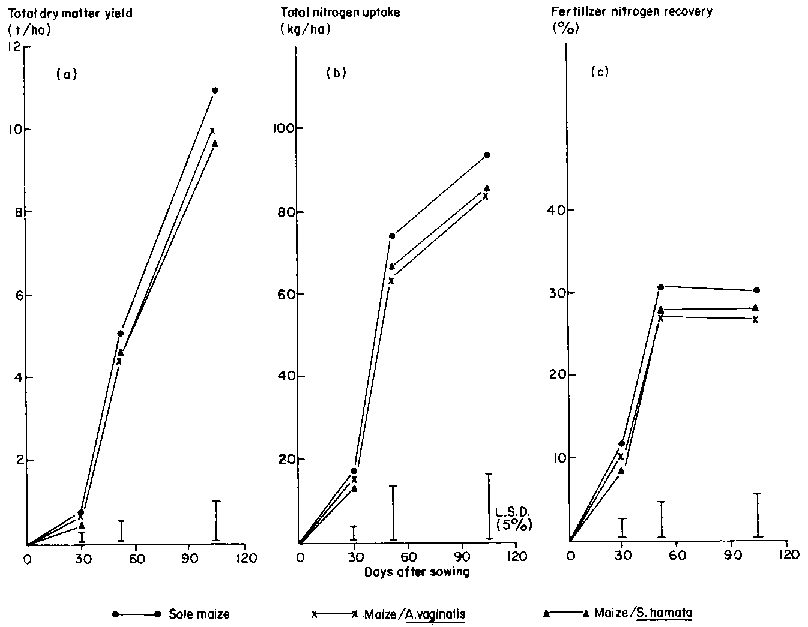
In Experiment 2 the presence of an understory of C. pascuorum significantly reduced maize grain yields and N uptake at all N rates (Figures 3 and 4). This twining legume severely shaded maize during grain filling, irrespective of N supply. The commencement of climbing by this legume in relation to stage of maize development is critical in intercropping (unpublished data).
Under conditions of high N supply the presence of S. hamata or A. vaginalis had very little effect on maize yields, but there were considerable yield reductions at lower N rates. Both legumes resulted in a 10 % reduction in maize grain size (data not shown). Table 2 shows that there were also significant effects of the presence of A. vaginalis and C. pascuorum on grain number. The lack of any effect of S. hamata on grain number is consistent with the observation that this species produced a much smaller proportion of its yield prior to maize anthesis than did the other legumes. It is during the period between cob initiation and anthesis that potential grain number of maize is determined. Production by A. vaginalis would have been underestimated as senescence was advanced and leaf loss high by the time of harvest, at maize maturity.
The 15N data confirm that, while recovery of fertilizer applied at the medium and low rates was low, there was competition for N between maize and the legumes (Table 3). Fertilizer recovery by the legume was highly variable and would have been underestimated due to leaf drop, particularly for A. vaginalis. The mean reduction in recovery of fertilizer N by maize due to intercropping was significant (P< 0.05), but there were no significant differences between legumes in this effect. Nevertheless, the trends shown in Table 3 are of importance. Competition for fertilizer N tended to be least from S. hamata, and this legume has been shown in Figures 3 and 4 to have least effect on maize grain yield and N uptake at all fertilizer rates. There was no interaction between legume species and N application rate.
Table 2. The effect of intercrop on maize grain number, Experiment 2.
|
Cropping system |
Grain number (x 10-7/ha) |
|
Sole maize |
1.84 |
|
Maize/A. vaginalis |
1. 41 |
|
Maize/S. hamata |
1.73 |
|
Maize/C. pascuorum |
1. 20 |
|
LSD (5%) |
0.26 |
Table 3. Average recovery of 15N-labelled fertilizer nitrogen by maize and legume intercrop when applied at medium and low rates, Experiment 2.
|
Cropping system |
Recovery % |
|
|
Maize |
Legume |
|
|
Sole maize |
15.8 |
|
|
Maize/A. vaginalis |
7.5 |
2. 3 |
|
Maize/S. hamata |
11.1 |
4.4 |
|
Maize/C. pascuorum |
7.3 |
5. 4 |
|
LSD (5%) |
7,5 |
3.5 |
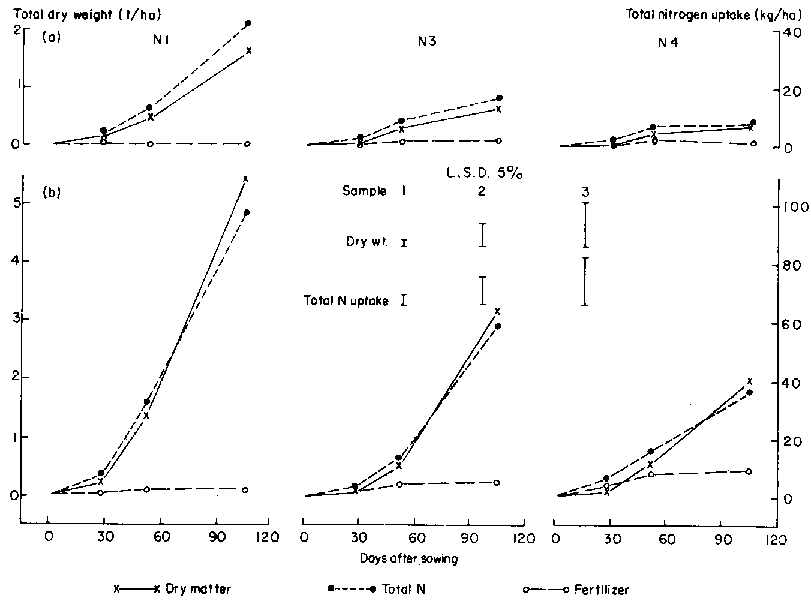
The 1983/84 season was very wet, and the plots in Experiment 3 were frequently waterlogged. Consequently, establishment and early growth of the legumes (particularly A. vaginalis) were poor. Average maize grain dry weight ranged from 2.0 t/ha for treatment N1 to 7.8 t/ha for N4. Figure 5 shows the effect of each of the legumes on total yield, N uptake and 15N fertilizer recovery averaged over N rates. Table 4 shows the effect on maize grain yield and components of yield. (There was no interaction between legume and N rate for any measured characteristic.) Intercropping had a consistent deleterious effect on maize performance, but any competitive effects were small. Uptake of fertilizer N was very rapid; even that applied 35 days after sowing was taken up by the time of silking, and N uptake during grain filling was therefore from non-fertilizer soil sources. The growth and N uptake of each of the legumes at fertilizer rates where 15N was applied are shown in Figure 6. As in Experiment 2, production by S. hamata was considerably better than that of A. vaginalis, but, as expected from Figure 5, fertilizer N use by these legumes was low.
Table 4. Effect of intercropping on maize yield and components of yield averaged across nitrogen treatments, Experiment 3.
|
Cropping system |
Grain yield |
Grain number |
1000 grain wt |
|
/ha |
|||
|
(kg/ha) |
(x 10-7) |
(g) |
|
|
Sole maize |
4974 |
2.02 |
241 |
|
Maize/A. vaginalis |
4377 |
1.76 |
243 |
|
Maize/S. hamata |
4345 |
1.76 |
240 |
|
LSD (5%) |
659 |
0.24 |
11 |
In contrast to results from Experiment 1, seed production from both legumes was good (Figure 7) even though shading by the maize was severe at high rates of N application (Table 5). This experiment received supplementary irrigation so that water stress was prevented before maize maturity. The CORNF model showed approximately 70 mm of available water in the root zone at maize harvest and the improved moisture availability at this time compared with Experiment 1 might have been responsible for the better legume seed yields. There is a clear relationship between dry-matter yield and seed yield of A. vaginalis but less so for S. hamata. While the former matured at the same time as the maize, the latter would have continued to grow after maize senescence while moisture was available, and its seed yield would have been heavily dependent on dry-matter production after maize maturity.
Table 5. Effect of nitrogen fertilizer rate on proportion of incident radiation transmitted to the top of the legume understory during early maize grain filling, Experiment 3.
|
Cropping system |
Proportion of incident radiation transmitted |
|||
|
N1 |
N2 |
N3 |
N4 |
|
|
Maize/A. vaginalis |
0.59 |
0.45 |
0.34 |
0.18 |
|
Maize/S. hamata |
0.65 |
0.40 |
0.35 |
0.17 |
For a given yield of legume N. the extent of competition for fertilizer and soil mineral N will depend on the proportion of N that the legume fixes from the atomosphere. The isotope dilution technique can be used to measure N fixation. However, Figures 5 and 6 show that there was no 15N available during grain filling in Experiment 3 and therefore probably not in Experiment 2 either, and that growth and N uptake patterns of the maize and associated legumes were different. The data on legume 15N uptake at earlier harvests was highly variable and complicated by poor legume establishment and poor early growth compared with that of the maize, and no attempt has been made to calculate fixation by this method (Witty, 1983). Nitrogen fixation may also be estimated by comparing N yield from a non-fixing system with that from a fixing system (the difference method). This calculation has not been done for A. vaginalis because of the large N loss due to leaf drop, but data are presented in Table 6 for the other legumes in Experiments 2 and 3.
Table 6. Estimates by the difference method of the percentage of nitrogen derived from fixation by S. hamata and C. pascuorum in Experiments 2 and 3.
|
(a) Experiment 2 |
(b) Experiment 3 |
|||
|
N rate |
Legume |
N rate |
S. hamata |
|
|
S. hamata |
C. pascuorum |
|||
|
Low |
91 |
74 |
N1 |
87 |
|
Medium |
69 |
68 |
N2 |
92 |
|
High |
72 |
60 |
N3 |
109 |
|
|
|
|
N4 |
40 |
Rennie (1984) found that the difference method underestimated fixation particularly when soil N availability was high, and that it was less precise than the isotope dilution method, 60 one must be careful in drawing conclusions from data of this nature. It appears however, that fixation by these forage legumes is only severely restricted by mineral N when supply of mineral N is high. Shading can also reduce N fixation in intercropping systems (Wahua and Miller 1978; Nambiar et al, 1983). Both nodule number and specific nodule activity can be affected, but the data from these experiments do not allow a distinction to be made between the direct effect of N supply on fixation and an indirect one through increased shading resulting from improved maize growth.
The small reductions in maize yield due to the presence of these forage legume intercrops in Experiment 3, compared with the large reductions in Experiments 1 and 2, cannot be easily explained in terms of legume N fixation. The timing of the onset of competition may be critical. It has often been shown (Hanway, 1962; Allison, 1984), and it is clear from Experiment 3, that maize takes up most of its N prior to grain filling, and a large proportion of grain N results from redistribution within the plant. Competition for N commencing prior to grain filling, when demand is high and potential grain yield is determined, is therefore likely to have a greater effect on maize production that competition during grain filling. As well as helping to explain differences between the effects of A. vaginalis and S. hamata, this may explain the smaller competitive effect of the legumes in a season when their establishment and early growth were poor.
There is no evidence from field trials carried out at Katherine, in northern Australia, that forage legumes when intercropped with maize made any contribution to the N economy of the cereal. The effect of intercropping on maize grain yield varied with environmental conditions, but was most negative under conditions favouring establishment and early growth of the legume, particularly when N supply was low. Species differences were probably related to growth pattern and phenology.
We are indebted to the Rural Credits Development Fund and the Australian Centre for International Agricultural Research for financial assistance. The technical assistance of J. Lazdofskis and M. Nicholas is gratefully acknowledged.
Allison J C S. 1984. Aspects of nitrogen uptake and distribution in maize. Ann. Appl. Biol. 104:357-365.
Argel P J and Humphreys L R. 1983. Environmental effects on seed development and hardseededness in Stylosanthes hamata cv. Verano. II. Moisture supply and illuminance. Aust. J. Agric. Res. 34:271-277.
Clements R J. Williams R J. Grof B and Hacker J B. 1983. Centrosema. In: R L Burt, P P Rotar, J L Walker and M W Silvey, (eds.). The role of centrosema, desmodium, and stylosanthes in improving tropical pastures. Westview Press, Boulder, Colorado. pp. 69-96.
Enyi B A C. 1973. Effects of intercropping maize or sorghum with cowpeas, pigeon peas or beans. Exp. Agric. 9:83-90.
Hanway J J. 1962. Corn growth and composition in relation to soil fertility: II. Uptake of N. P. and K and their distribution in different plant parts during the growing season. Agron. J. 54:217-222.
IITA (International Institute of Tropical Agriculture). 1980. Annual Report for 1980. IITA, Ibadan, Nigeria.
Kurtz T. Melsted S W and Bray R H. 1952a. The importance of nitrogen and water in reducing competition between intercrops and corn. Agron. J. 44:13-17.
Kurtz T. Melsted S W. Bray R H and Breland H L. 1952b. Further trials with intercropping of corn in established sods. Soil Sci. Soc. Amer. Proc. 16:282-285.
McCown R L, Winter W H. Andrew M H. Jones R R and Peake the Late D C I. 1985. A preliminary evaluation of legume ley farming in the Australian semi-arid tropics. In: Potentials of Forage Legumes in Farming Systems of Sub-Saharan Africa. ILCA (in press).
Martin T J and Torssell B W R. 1974. Buffalo clover (Alysicarpus vaginalis (L.) DC): A pasture legume in northern Australia. J. Aust. Inst. Agric. Sci. 40:232-4.
Nair K P P. Patel U K, Singh R P and Kaushik M K. 1979. Evaluation of legume intercropping in conservation of fertilizer nitrogen in maize culture. J. Agric. Sci. 93:189-194.
Nambiar P T C, Rao M R. Reddy M S. Floyd C N. Dart P J and Willey R W. 1983. Effect of intercropping on nodulation and N2 -fixation by groundnut. Exp. Agric. 19:79-86.
Rennie R J. 1984. Comparisons of N balance and 15N isotope dilution to quantify N2 fixation in field-grown legumes. Agron. J. 76:785-790.
Stapper M and Arkin G F. 1980. CORNF: A dynamic growth and development model for maize (Zea mays L.). Texas Agric. Expt. Sta., Blackland Res. Center, Temple, Texas.
Waghmare A B and Singh S P. 1984. Sorghum-legume intercropping and the effects of nitrogen fertilization. I. Yield and nitrogen uptake by crops. Exp. Agric. 20:251-259.
Wahua T A T. 1983. Nutrient uptake by intercropped maize and cowpeas and a concept of nutrient supplementation index (NSI). Exp. Agric. 19:263-275.
Wahua T A T and Miller D A. 1978. Effects of intercropping on soybean N2 fixation and plant composition on associated sorghum and soybeans. Agron. J. 70:292-295.
Williams J. Day K J. Isbell R F and Reddy S J. 1985. Soils and climate. In: R C Muchow, (ed.). Agro-research for the semi-arid tropics: north-west Australia. University of Queensland Press, St. Lucia. pp. 31-92.
Witty J F. 1983. Estimating N2 fixation in the field using 15N-labelled fertilizer: some problems and solutions. Soil Biol. Biochem. 15: 631-639.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}