![]()
![]()
![]()
Abstract
Introduction
Materials and methods
Results
Discussion
References
Ekow Akyeampong
Highlands Programme,
ILCA, P.O. Box 5689, Addis Ababa, Ethiopia
Traditionally, cowpea (Vigna unguiculata) has been used mainly as a grain crop in the semi-arid areas of sub-Saharan Africa. However, it could be grown as a forage crop or a dual-purpose crop that provides high protein grain for human consumption and crop residues of high nutritive value for animal feed. Drought is a constraint to crop production in the semi-arid regions, the principal area of cultivation of cowpea. The effect of drought stress on plant water status, leaf area and seed yield were examined in a number of experiments. Predawn leaf water potential (PLWP) remained fairly constant (at the vegetative stage) or declined gradually (at the flowering or pod-filling stage) as duration of stress was prolonged. The difference between PLWP of stressed and unstressed plants was small, although PLWP was always lower in stressed plants. Midday leaf water potentials were much lower in the drought-stressed plants than in the unstressed plants and the differences increased as duration of stress was extended. When plants were drought-stressed at the vegetative stage, the production of new leaves and expansion of leaves ceased. Leaf area therefore remained constant. Upon termination of stress at the vegetative stage, leaf growth resumed. At maturity, leaf area and seed yield of stressed plants were not significantly different from unstressed plants. Although leaf production and expansion ceased when plants were stressed at the flowering or pod-filling stage, leaf abscission was more important in determining total leaf area per plant. Abscission commenced with basal mature leaves. Leaf areas of plants under drought stress at flowering or pod-filling did not recover after re-watering and seed yield was lower than that of unstressed plants. In confusion, cowpea can tolerate drought stress at the vegetative stage and recover when water is available at the reproductive stage to produce DM and seed yields equivalent to unstressed plants. Drought stress at the flowering or pod-filling stage reduced yields.
Cowpea (Vigna uguiculata) produces seed of high protein content (20% to 40%) for human consumption, and can also be used to increase soil fertility. Estimates of the amout of N fixed biologically by cowpea range from 73 to 354 kg N/ha per year (FAO, 1984), some of which may be available to succeeding crops. Cowpea may be also be grown as a forage legume to provide fodder of higher quality than cereals or forage grasses.
All three benefits may be gained if cowpea is used as a dual-purpose crop for seed and crop residues. However, this would require that pods are harvested green or only partly ripe, because leaf shedding is very pronounced during the later stages of pod ripening, leaving only stems at harvest when pods are completely ripened and dry. As a forage legume, Cowpea could provide livestock feed in the semi-arid areas where it is mainly cultivated and which are the major livestock production areas in sub-Saharan Africa. The semiarid areas are characterised by low and unreliable rainfall, and hence are prone to drought. Water deficits are the main constraint on crop production in these areas (Begg and Turner, 1976).
Many aspects of plant growth are affected by drought stress (Hsiao, 1973), including leaf expansion, which is reduced due the sensitivity of cell growth to water stress. Water stress also reduces leaf production and promotes senescence and abscission (Karamanos, 1980), resulting in decreased total leaf area per plant. Reduction in leaf area reduces crop growth and thus biomass production. Seed production, which is positively correlated with leaf area (Rawson and Turner, 1982) may also be reduced by leaf area reductions induced by drought stress.
For cowpea to be used as a forage or dual-purpose crop in the semi-arid areas, the effect of drought stress on leaf area, and the relationship between leaf area and seed yield, must be understood. Changes in plant water status due to drought stress also need to be investigated, since the growth of a leaf is intimately linked with its water potential.
Since drought may occur at any stage of growth, cowpea was drought-stressed at the vegetative, flowering or pod-filling stages to assess responses of leaf water potential, leaf area and seed yield to water stress.
Seeds of cowpea were sown in 1:1 mixture of Cornell Mix and sterilised field soil (fine loamy, mixed, non-acid aquic udifluvents) in 9-litre plastic pots in greenhouses at Cornell University, Ithaca, N.Y., USA. Upon germination, seedlings were thinned to one per pot. Once each week, liquid fertilizer was applied and plants were sprayed against pests and diseases. A 12-hour photoperiod (0600 to 1800 hours) was established using high intensity lamps. Temperature was maintained at 29°C during the day and 21° C at night. Plants were watered daily during the juvenile stage and twice daily when mature. Drought stress was induced by complete cessation of watering.
Leaf water potentials were measured on young fully expanded leaves using a pressure chamber (Soil Moisture Equipment Corp., Santa Barbara, Cal, USA). To determine the leaf area, lamina length (L) and maximum width (W) of each leaflet was measured with a ruler to the nearest millimeter. Leaf area was then calculated as the product of the length, width and a factor (F) determined for each type of leaflet.
To obtain F. several leaflets were measured by a leaf area meter (Li-3000, Lambda Instruments Corp., Lincoln, Nebraska). Each leaflet area was divided by the product of its length and width. The quotients were then averaged for each type of leaflet, giving F values of 0.67 and 0.71 for for variety TVu 4552; 0.69 and 0.72 for TVu 4954; and 0.49 and 0.54 for TVu 1954-01E for the terminal and the two basal leaflets, respectively. Seed yield and components of yield were determined on plants from which leaves had not been sampled for leaf water potential measurements. When pods started to ripen, watering was curtailed in all treatments to allow pods to ripen and dry. Pods were then harvested, threshed by hand, dried at 55° C for 14 days and weighed.
Treatments
In 1982, TVu 4552 was used in three experiments. In the first experiment, plants were stressed at the vegetative stage for 14 days. In the second experiment, watering was stopped for 12 days, beginning 3 days after the first flower opened. Drought stress was imposed at the pod-filling stage in the third experiment; plants were not watered for 12 days, beginning 12 days after the first flower opened. Treatments for 1983 experiments, in which three varieties were used, are shown in Table 1.
Vegetative stage
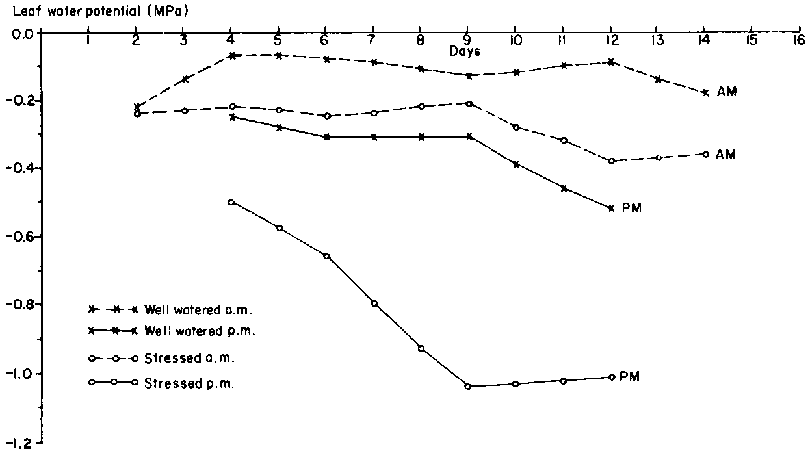
1982: Two days after cessation of watering, the predawn leaf water potential (PLWP) of the drought-stressed plants (-0.22 MPa) was approximately equal to that of unstressed plants (Figure 1). Subsequently, the PLWP of unstressed plants was about -0.1 MPa throughout the 14 days, and the midday leaf water potential (MLWP) declined from -0.25 MPa on day 4 to-0.52 MPa on day 12. Predawn leaf water potential of the drought-stressed plants remained fairly constant at about -0.22 MPa until the ninth day, after which it declined to -0.37 MPa 12 days after the last irrigation. However, MLWP of drought-stressed plants decreased linearly from -0.50 MPa on day 4 to -1.0 MPa on day 9.
Table 1. Experimental treatments.
|
Growth stage |
TVu 1954-01E |
TVu 4954 |
TVu 4552 |
|||
|
Perioda |
PLWPb |
Period |
PLWP |
Period |
PLWP |
|
|
Early veg. |
23-34 |
-0.47±0.05 |
23-34 |
-0.52±0.02 |
23-34 |
-0.49±0.02 |
|
Late veg. |
30-39 |
-0.72±0.03 |
30-39 |
-0.57±0.53 |
30-39 |
-0.59±0.02 |
|
Veg. |
23-39 |
-0.54±0.05 |
23-39 |
-0.63±0.06 |
23-39 |
-0.73±0.02 |
|
Control |
0 |
-0.25±0.01 |
0 |
-0.23±0.01 |
0 |
-0.26±0.01 |
a. Period without water, days after planting.
b. Predawn leaf water potential, MPa.
On average, the difference in PLWP between the drought-stressed and unstressed leaves was less than -0.15 MPa for the first 10 days in the 1982 experiment. Differences between MLWPs of stressed and unstressed plants increased with time and were larger than differences between the predawn values. Differences between predawn and midday values of leaf water potential were larger in drought-stressed than in unstressed plants.
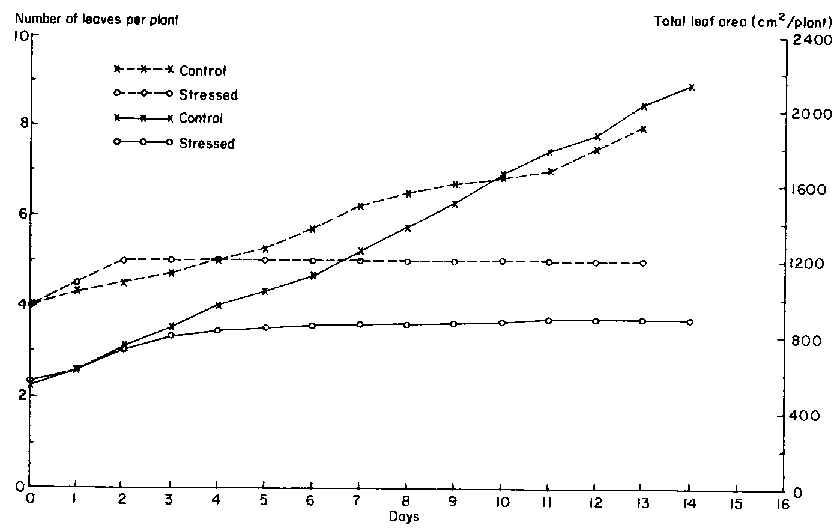
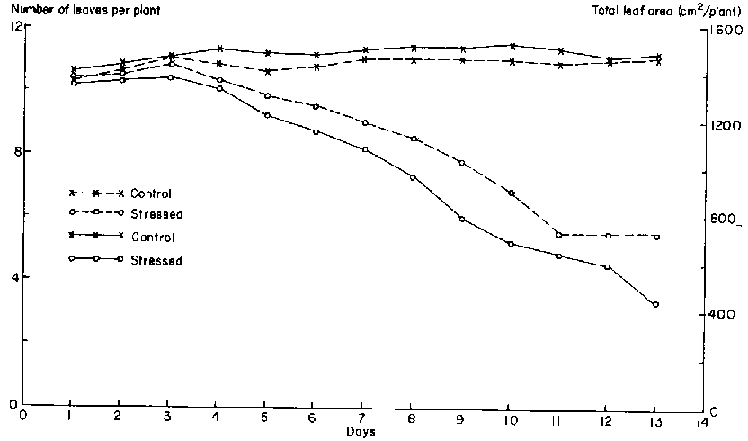
Total leaf area of unstressed plants increased linearly throughout the data collection period because the number of leaves per plant increased from about 4 to 9 (Figure 2). In contrast, leaf growth slowed and eventually stopped in drought-stressed plants. As early as 4 days after irrigation was withheld, the increase in total leaf area of drought-stressed leaves decreased. After day 3, total leaf area of the drought-stressed plants stopped increasing; at the same time, MLWP was considerably lower than in unstressed plants.
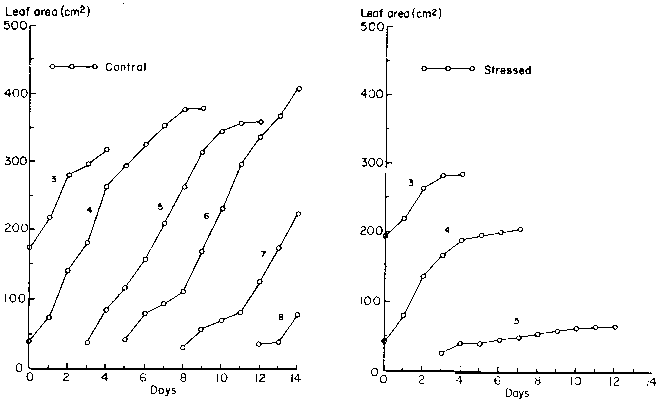
The decline in total leaf area was due to cessation of the initiation of new leaves (Figure 2) and a decrease in the expansion of individual leaves. When the drought stress was initiated (21 days after planting), plants were at the fourth or fifth trifoliate leaf stage. Drought stress totally inhibited new leaf initiation after 3 days, resulting in an average of 5 leaves per plant. Figure 3 illustrates the growth patterns of all leaves that were still expanding during the data collection period for both sets of plants. The final area of the drought-stressed leaf 3 was only slightly smaller than the unstressed leaf. The effect of the drought stress was more severe on the growth of leaves 4 and 5. Leaf 5 barely expanded after the initial unfolding; reduction in its area was at least 70%.
1983: Three separate experiments were conducted using varieties TVu 4954, TVu 1954-01E and TVu 4552. Plants were subjected to drought-stress at the early vegetative stage
(EV), late vegetative stage (LV) or throughout the vegetative stage (V). Leaf areas were measured at the end of the vegetative stage and at maturity (Table 2).
Table 2. The effect of drought stress at the vegetative stage on leaf area and seed yield of TVu 4552, TVu 1954-01E and TVu 4954. Values are mean + one standard error of three replicates.
|
Growth stage |
Total leaf area (cm/plant) |
Number of leaves/plant |
Seed yield (g/plant) |
||
|
Veg. |
Maturity |
Veg. |
Maturity |
||
|
TVu 4552 |
|
||||
|
Early Veg. |
1394±101 |
2158±270 |
7.7±0.3 |
13.3±1.3 |
14.11±1.60 |
|
Late Veg. |
1615±162 |
2460±483 |
8.7±0.7 |
13.3±1.9 |
14.89±0.90 |
|
Veg. |
1064± 62 |
2258±132 |
6.3±0.7 |
15.7±0.9 |
13.47±1.90 |
|
Control |
2573±198 |
3015±250 |
13.7±1.7 |
17.0±1.2 |
18.27±1.67 |
|
LSD |
533 |
ns |
2.9 |
ns |
ns |
|
TVu 4954 |
|
||||
|
Early Veg. |
1588±204 |
2181±119 |
13.3±0.7 |
18.0±0.50 |
11.22±0.50 |
|
Late Veg. |
1676± 82 |
1815± 73 |
14.7±1.3 |
16.3±1.22 |
12.22±0.38 |
|
Veg. |
1064± 62 |
2046±137 |
12.3±0.9 |
18.0±0.50 |
8.07±0.71 |
|
Control |
2334±235 |
2315±204 |
20.0±1.7 |
21.3±2.2 |
12.92±0.69 |
|
LSD |
650 |
ns |
3.9 |
ns |
2.16 |
|
TVu 1954-01E |
|
||||
|
Early Veg. |
1066± 63 |
2359±414 |
18.3±1.2 |
33.7±3.8 |
14.70±2.55 |
|
Late Veg. |
1630± 93 |
2657± 60 |
23.7±1.2 |
37.0±1.2 |
16.32±1.08 |
|
Veg. |
633±106 |
2552±403 |
14.7±0.3 |
35.3±3.4 |
13.36±0.77 |
|
Control |
2169± 77 |
3201±337 |
31.0±1.5 |
43.0±1.5 |
16.39±3.27 |
|
LSD |
290 |
ns |
4.2 |
ns |
ns |
At the end of the vegetative stage, leaf areas of the drought-stressed plants were significantly smaller than those of unstressed plants (Table 2). Upon re-watering, drought-stressed plants resumed growth. At maturity, differences in leaf area between the drought-stressed and unstressed treatments was not significant.
In TVu 4552, seed yields of all drought-stressed treatments were not different from each other and were lower than the control. However, this difference was not statistically siginificant. The regression of leaf areas at the end of the vegetative stage and seed yield at maturity was significant (r = 0.77, P<0.01) and was described by the equation: Yield = 9.2 + 0.004 (Leaf Area).
The seed yield of TVu 4954 plants subjected to drought-stress throughout the vegetative period (treatment V) was significantly lower than that of unstressed plants. The regression of leaf areas at the end of the vegetative stage on seed yield was: Yield = 5.61 + 0.003 (Leaf Area), and was significant (r = 0.82, P<0.01). The regression of leaf areas at maturity on seed yield was not significant. For TVu 1954-01E, neither the leaf area at the end of the vegetative stage nor at maturity was significantly correlated with seed yield.
Flowering stage
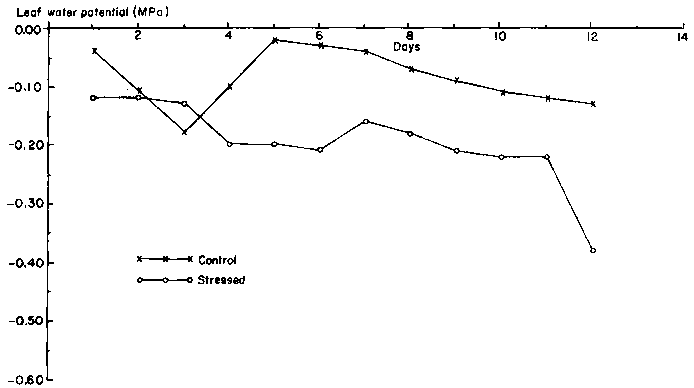
For the first 11 days of the drought period at the flowering stage, PLWP of stressed plants remained within 0.15 MPa of that of unstressed plants (Figure 4). During this period, leaf water potential of the drought-stressed plants declined by only 0.1 MPa, from -0.12 MPa to -0.22 MPa.
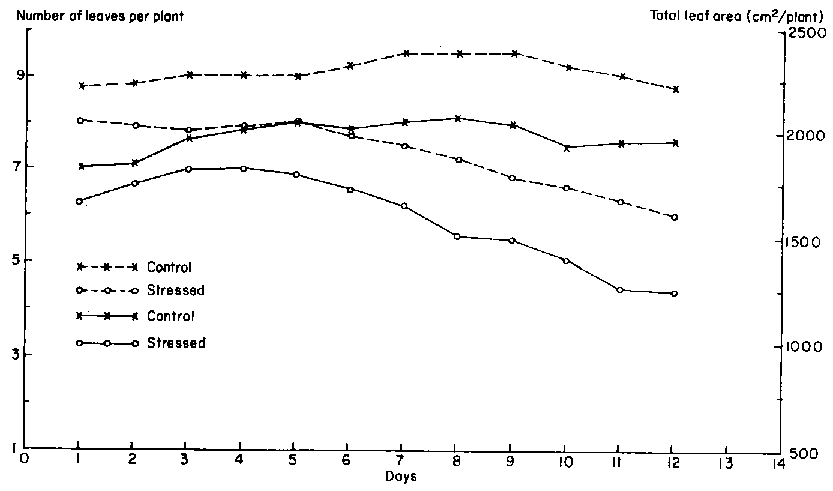
During the flowering stage, the leaf area of unstressed plants remained fairly constant. Three days after the imposition of drought stress at the flowering stage, leaf area increment began to diminish (Figure 5). After day 4, total leaf area began to decline. Over the 12-day period, total leaf area of the drought-stressed plant declined by about 40%, mainly due to loss of leaves. Changes in the total leaf area of the drought-stressed plants were the result of changes in the number of leaves per plant and in leaf expansion. The treatments were imposed 2 days after the first flower opened, when plants were at the 13- to 15-leaf stage. Due to injury from pesticide application at the vegetative stage, the lowest five or six leaves of each plant had abscissed. Most of the unstressed plants added one or two new leaves during the course of the experiment, but also lost mature basal leaves, which left an average of nine leaves per plant. Drought-stressed plants also exserted a few new leaves but could not retain the older ones after 4 days of drought-stress. On average, only six leaves remained per plant at the end of the drought phase.
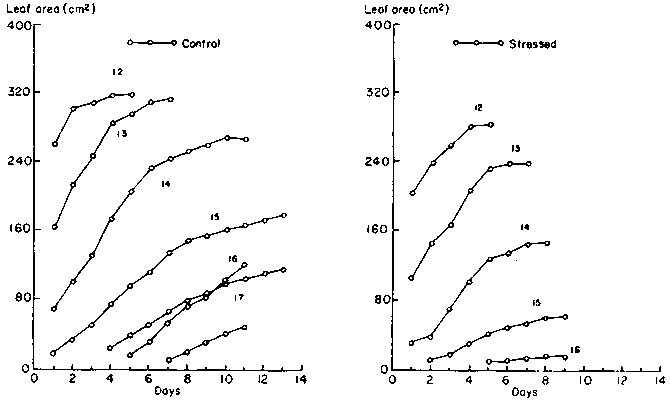
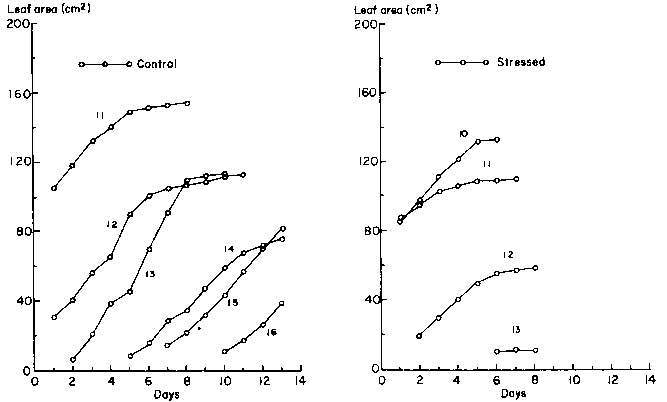
In addition to the loss of leaves, the expansion of individual leaves slowed and stopped in stressed plants. Growth patterns of individual leaves at the flowering stage are illustrated in Figure 6. The oldest leaf that was still expanding at the beginning of the treatments was that at node 12. Drought stress inhibited the growth of all leaves and caused delays in the exsertion of leaves 15 and 16.
Drought stress significantly reduced seed yield due to a reduction in the number of pods per plant (Table 3). Individual seed weight was significantly higher in the drought-stressed treatment but did not compensate for the loss in pods. Leaf area measured at the end of the drought-stress period was highly correlated with seed yield. The regression equation was:
Yield = 2.9 + 0.008 (Leaf Area) (r = 0.90, P<0.01)
Table 3. The effect of drought stress at the flowering or pod-filling stage on leaf area and seed yield of TVu 4552. Values are mean ± one standard error of four replicates.
|
Treatment |
Leaf area(cm2/ plant) |
Seed yield(g/ plant) |
Pods per plant |
Seeds per pod |
100 seed wt. (g) |
|
Flowering stage |
|||||
|
Control |
1956±280 |
19.4+2.6 |
15.8+0.8 |
9.7+0.9 |
12.8+2.3 |
|
Stress |
1167±101 |
13.0+1.8 |
11.3+0.6 |
8.1+0.8 |
14.8+3.6 |
|
LSD (0.05) |
649 (0.10) |
4.5 |
2.8 |
NS |
1.4 |
|
Pod-filling stage |
|||||
|
Control |
1478±180 |
5.6±1.2 |
5.5±0.9 |
8.0±0.9 |
12.7±1.1 |
|
stress |
748±184 |
4.0±0.7 |
3.3±0.3 |
7.0±0.2 |
14.9±0.5 |
Pod-filling stage
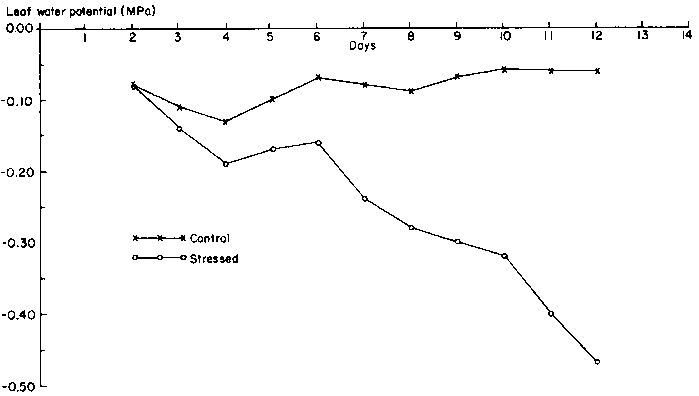
Predawn leaf water potential of the drought-stressed plants declined gradually to -0.48 MPa on the 12th day, when stress was terminated (Figure 7). However, it was not until the 7th day of drought that the difference between stressed and unstressed leaves was larger than 0.15 MPa. Unstressed leaves had PLWPs of about -0.10 MPa.
Total leaf area started to decline 3 days after the cessation of watering at the pod-filling stage, and continued to decline until the last day of the drought period (Figure 8) because of leaf abscission. Total leaf area of unstressed plants remained constant.
Approximately 10 leaves were present on each plant at the beginning of the treatments at the pod-filling stage (Figure 8). This number increased to about 11 in both treatments. Leaf senescence and abscission reduced the number of leaves on drought-stressed plants to about six. Although unstressed plants initiated a few new leaves, basal mature leaves abscissed. Thus, a constant number of leaves was maintained.
The new leaves expanded to smaller final sizes, as indicated by the decreasing sizes of the higher numbered leaves (Figure 9). One drought-stressed plant did not put out any new leaves and all its leaves ceased expanding immediately after the drought stress was imposed. Drought-stressed leaves were smaller than the unstressed leaves.
Leaf abscission was more important than cessation of leaf expansion in determining final leaf area. Total leaf area of drought-stressed plants was only 50% of that of unstressed plants (Table 3). Seed yield and number of pods per plant, even of unstressed plants, were extremely low. The regression of leaf area on seed yield was significant (P < 0.10), with a correlation coefficient r = 0.64.
During drought stress, cowpea maintained fairly constant PLWP at a level not much lower than that of unstressed plants. Turk and Hall (1980) also showed small differences between the leaf water potential of stressed and unstressed cowpea even after 43 days without irrigation. Drought-stressed plants occasionally had higher leaf water potentials than well-watered plants.
This behaviour of cowpea is unlike crops such as soya bean (Wien et al, 1979; Villalobos-Rodriquez and Shibles, 1985) and maize (Lal et al, 1979), in which large differences in leaf water potential were found between drought-stressed and unstressed leaves. The constancy or slow decline in PLWP suggests that cowpea can conserve water. An explanation for this behaviour may be found in the changes in leaf growth that limited evaporative water loss (Akyeampong, 1985): Leaf exsertion, expansion, and abscission were sensitive to drought stress at the vegetative, flowering or pod-filling stage.
At the vegetative stage, drought stress initially reduced the rate of leaf expansion, followed by a cessation of new leaf production. Eventually, leaf area increment fell to zero. No leaves were lost, however. Stressed plants remained in a stunted state until they were re-watered (Warrag and Hall, 1985). In addition to the cessation of expansion of leaves and inhibition of production of new leaves, drought stress at the flowering or pod-filling stage caused the senescence and abscission of mature basal leaves. Only mild stress is required for the abscission of mature leaves (McMichael et al, 1973). At the vegetative stage, no leaves were lost, even when the PLWP was -0.40 MPa, which was lower than the PLWP on day 4 of the flowering (-0.2 MPa) or pod-filling (-0.18 MPa) treatments when pods were lost. The number of leaves per plant, and hence the total leaf area per plant, was reduced when plants were stressed at the flowering or pod-filling stage.
Drought stress thus reduced evaporative surface area, which would reduce transpiration. Extensive leaf movements to minimise the interception of radiation (Shackel and Hall, 1979) and the reduction in stomatal conductance (Akyeampong, 1985) also reduce transpiration in cowpea under drought stress (Akyeampong, 1985; Hall and Schulze, 1980). Other suggested mechanisms that cowpea uses to maintain plant water status are: (a) increased root density and depth during stress to exploit a larger volume of soil, (b) a decrease in plant resistance to water movement as the level of stress increases, to allow for the replenishment of lost water (Wien et al, 1979) and (c) mechanisms which result in only slow extraction of soil water as soil begins to dry (Turk and Hall, 1980). Such mechanism ensure the survival of the crop during drought.
The cessation of leaf expansion-at a high leaf water potential indicates that cowpea leaves are acutely sensitive to drought stress. Leaf expansion was probably influenced more by daytime than by predawn leaf water potential. Midday leaf water potential declined progressively with time. The reduced moisture status of the leaves deprived the cells of the forces for expansion and thus slowed leaf expansion. Cell division, considered to be less sensitive to drought stress than cell expansion (Hsiao, 1973), may have been inhibited later as stress increased.
Equally, if not more, important from the point of view of the use of cowpea as either a forage or grain crop, were observations of the growth of cowpea subsequent to re-watering. After re-watering, plants that were stressed at the vegetative stage resumed growth and leaf area recovered completely, providing large amounts of vegetative material that could be used as feed. Leaf growth during the recovery phase was not at the expense of seed-filling (Fischer and Hagin, 1965), and drought-stressed plants yielded as well as those that had been well-watered throughout. Cowpea recovers from drought stress at the vegetative stage, provided that favourable climatic conditions prevail during the post-stress period (Shouse et al., 1981).
In contrast, the detrimental effect of drought stress at the flowering or pod-filling stage was not alleviated by re-watering. When the plants were re-watered at the flowering stage, reproductive activity resumed, but most of the new pods failed to reach maturity due to inadequate resources, the result of excessive leaf loss. Presumably, photosynthate was used to support leaf growth in this more-or-less indeterminate variety, to the detriment of pod-filling (Fischer and Hagin, 1965).
In contrast to drought-stressed plants, growth of leaves on the unstressed plants continued through the pod-filling stage, but total leaf area did not expand much during the flowering and pod-filling stages due to three factors. Firstly, only a few leaves were exserted after flowering. Secondly, some of the mature leaves near the base of the plants were lost. Finally, although duration of the expansion of unstressed leaves was about equal at all stages of growth, individual leaf expansion rate was low during the flowering and pod-filling stages. Hence new leaves were smaller, adding little to leaf area.
These results have important implications for the management of planting (and irrigation, where feasible) of cowpea. To avoid severe yield (seed and total biomass) losses, planting should be done at such a time that the critical stages (flowering and pod-filling) would coincide with the period when water is most available.
Akyeampong Ekow. 1985. Seed yield, water use, and water use efficiency of cowpea in response to drought stress at different developmental stages. PhD Thesis, Cornell University.
Begg J E and N C Turner. 1976. Crop Water deficits. Adv. Agron. 28:161-217.
FAO (Food and Agriculture organization). 1984. Legume inoculants and their use. FAO, Rome.
Fischer R A and R M Hagan. 1965. Plant water relations, irrigation management and crop yield. Exp. Agric. 1:161-177.
Hall A E and E-D Schulze. 1980. Stomatal responses and possible interrelation between stomata! effects on transpiration and CO2 assimilation. Plant, Cell Environ. 3:467-474.
Hsiao T C. 1973. Plant responses to water stress. Ann. Rev. Plant Physiol. 24:519-570.
Karamanos A S. 1980. Water stress and leaf growth of field beans (Vicia faba) in the field: Leaf number and total area. Ann. Bot. 42:1393-1402.
Lal R. P R Maurya and S O Yeboah. 1978. Effects of no-tillage and ploughing on efficiency of water use in maize and cowpea. Exp. Agric. 14:113-120.
McMichael B L, W R Jordan and R D Powell. 1973. Abscission process in cotton: induction by plant water deficits. Agron. J. 65:202-204.
Rawson H M and N C Turner. 1982. Recovery -from water stress in five sunflower (Helianthus annus L.) cultivars. I.
Effect of timing of water application on leaf area and seed production. Aust. J. Plant Physiol. 9: 437-448.
Shackel K A and A E Hall. 1979. Reversible leaflet movements in relation to drought adaptation of cowpeas [Vigna unguiculata (L.) Walp.]. Aust. J. Plant Physiol. 6:265276.
Shouse P S. S Darsberg, W A Jury and H Stolzy. 1981. Water deficit effects on water potential, yield and water use of cowpeas. Agron J. 73:333-336.
Turk K J and A E Hall. 1980. Drought adaptation of cowpea. II. Influence of drought on plant water status and relations with seed yield. Agron. J. 72:421-427.
Villalobos-Ridriquez E and R. Shibles. 1985. Response of determinate and indeterminate tropical soybean cultivars to water stress. Field Crops Res. 10: 269-281.
Warrag M O A and A E Hall. 1984. Reproductive responses of cowpea (Vigna unguiculata (L) Walp) to heat stress. II. Responses to night air temperature. Field Crops Res. 8:17-33.
Wien H C, E J Littleton and A Ayanaba. 1979. Drought stress of cowpea and soybean under tropical conditions. In Mussel H and Staples R C (eds). Stress physiology in crop plants. Wiley Interscience, New York. pp. 283-310.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}