![]()
![]()
![]()
E.K. Odhuba
Animal Production Research Department
Kenya Agricultural Research Institute (KARI), Muguga
P.O. Box 30148, Nairobi, Kenya
Introduction
Materials and methods
Results
Discussion
Conclusions
Acknowledgement
References
Abstract
A study of breed and environmental factors affecting calving interval of range-fed beef cattle was conducted at Athi River Ranch in the Kenyan rangelands. In experiment 1, conception period, rainfall and body weight changes prior to conception were evaluated from a total of 308 calving intervals. In experiment 2, 1985 in-calf females were supplemented during the dry season in 1978, 1979 and 1980. In experiment 3, 122 cows were allowed to wean their calves at various ages at the end of September, October and November/December in 1977, 1978 and 1979.
Short mean calving intervals were associated with conception between November and January. Increased rainfall during the month prior to conception was correlated with a linear and highly significant (P<0.01) reduction in calving interval depending on conception period. Year effects were highly significant (P<0.005). The target joining weight was around 318 kg. Above the target weight, fertility became a function of the absolute body weight such that heavier cows at joining were more fertile (P<0.01) than lighter ones. Below the target weight, fertility was associated with females that gained weight or suffered less body weight loss during the month prior to conception. Previously, dry cows tended to gain weight prior to conception to the detriment of fertility (P<0.005). Effect of calving month was highly significant (P<0.005) depending on age of breeding females. Influence of dry season supplementation was significant but was modified by a number of factors such as age, year and previous parity. There was no significant difference in fertility of cows that were supplemented with energy or energy plus urea before calving. Generally, cows that were supplemented after calving had a shorter mean calving interval. Effects of weaning period, month and year were marginally significant (P<0.05). Fertility improved as the dam's age increased from three years onwards but declined after about nine to ten years. Effects of previous parity and breed were marginal (P<0.05) depending on conception period. With supplementation, Boran crosses had a significantly (P<0.01) shorter mean calving interval than the pure Boran females. In general, effects of breed were minor compared to environmental factors especially those associated with conception period.
More than 50% of the 8.7 million beef cattle in Kenya are reared in semi-arid to arid areas or rangelands (Livestock Development Division, 1983). The overall offtake of beef is not adequate to meet domestic demand and at the same time sustain an export market. There is need, therefore, to increase not only efficiency of production but also to multiply the beef population quickly. The latter objective calls for a higher level of fertility. Not only does low fertility reduce productivity but it also limits improvement of the herd genetically. For instance, when calving rate is low, nearly all replacement heifers must be used to maintain the herd numbers, and this curtails selection intensity thereby lowering the rate of genetic improvement. Besides, a decline in fertility would result in a compensatory increase in the number of unproductive and aged cows with adverse consequences of overstocking leading to environmental degradation which is often irreversible.
Fertility can be evaluated in terms of calving rate, i.e. the number of calves born in relation to the number of breeding females exposed to the bulls per annum. This measurement is ideal for survey studies. For selection purposes, individual cow data are required for which the parameter evaluated is the calving interval, i.e. the period between two successive parturitions. Although several factors may influence fertility, there is overwhelming evidence that inadequate nutrition is the most important single factor mitigating reproductive efficiency in beef cattle (Topps, 1977). This observation is particularly relevant to tropical rangelands where pronounced seasonal changes in rainfall (French, 1957) contribute to a very precarious feed supply. Topps (1977) has associated seasonal changes in feed supply with corresponding changes in body weight of grazing beef cattle with adverse consequences on reproductive performance. The objective of this study was to characterise the influence of body weight changes and related factors that are associated directly or indirectly with the nutritional status of beef cows grazing natural pasture on calving interval.
Site
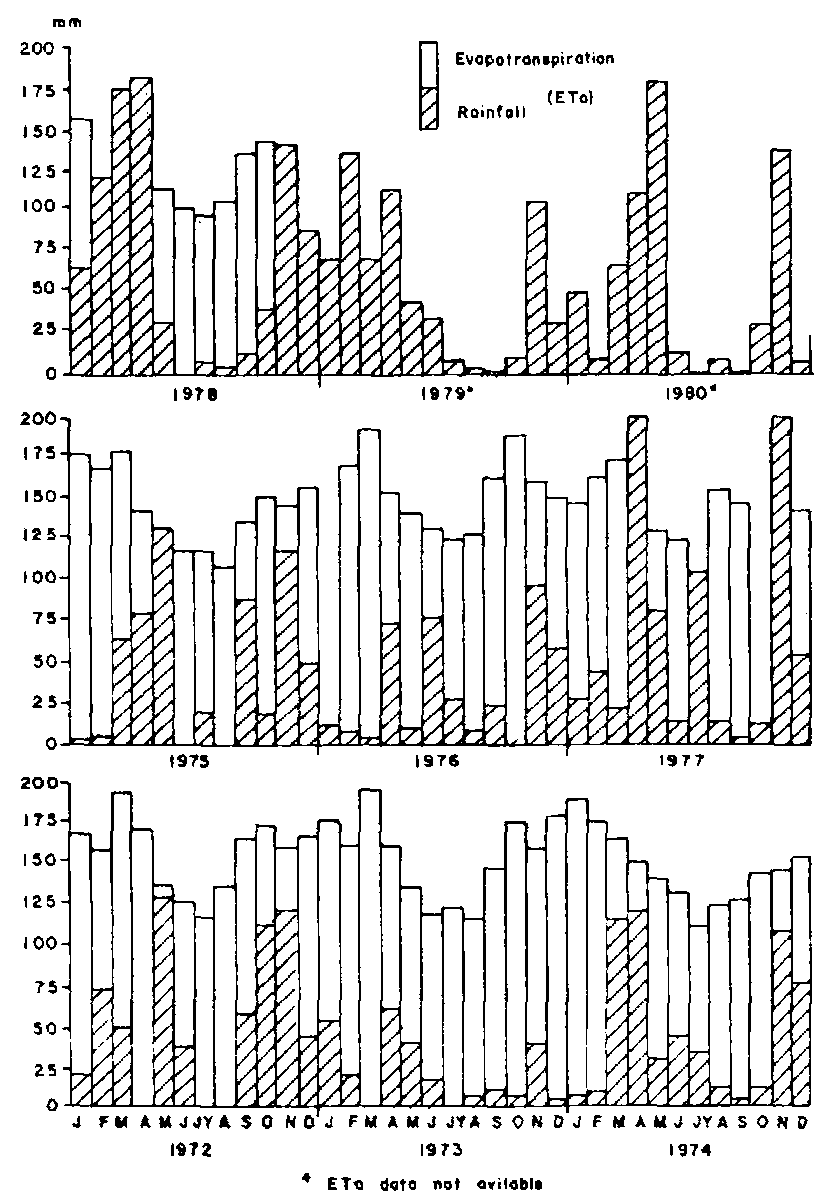
The study was conducted at KARI's field station near Athi River. The ranch lies in a semi-arid zone with a bimodally distributed rainfall averaging 565 mm per year (Figure 1). The main grass species is Themeda triandra Forsk on gently undulating mosaic of ridges and shallow valleys of black clay loams (Gethin-Jones and Scott, 1955; Ledger et al., 1969).
Breeding Females
The genotypes were the Boran, small East African Shorthorn Zebu (EASZ) and their F1 progeny from Hereford bulls. All animals were run as one herd and were grazed on natural pasture. Routine measures to control ticks and vaccinations against foot-and-mouth, contagious abortion, rinderpest, blackquarter and anthrax were undertaken. Random mating was followed throughout 1973. From 1974 until 1976, mating was confined between June and August, but this was subsequently brought forward between Nay and July. Heifers were first mated after attaining a liveweight of 250 kg.
Figure 1. Rainfall and evapotranspiration (mm) at Athi River Ranch.
Experiment 1: Evaluation of Breed and Environmental Factors Affecting calving Interval
Conception Period
Each calendar year was divided into six two-month periods to coincide as much as possible with the seasonal pattern of rainfall and temperature. Date of "second" conception was estimated from date of calving and assuming a mean gestation period of 283 days (Hutchison and Macfarlane, 1958). An appropriate conception period was then assigned to each calving interval.
Soil Moisture Index
Evaporative loss (E0) was estimated using McCulloch's (1965) Tables from data collected at the meteorological site at the ranch. E0 values were multiplied by a crop coefficient of 0.86 derived by Pereira and McCulloch (1962) to predict evapotranspiration (ET0) from a vegetative cover. A soil moisture index (SMI) was calculated from the equation:
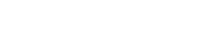
which, for the purposes of this study, gave an indication of soil water adequacy for pasture growth after taking into account the biggest water loss (ET0. Indices were calculated for each calving interval for the period of one month (SMI1) and one to two months (SMI2) prior to "second" conception.
Grouping of Females
Arising from variation in the breeding season, females were divided into five groups depending on the period taken after "first" calving before joining with the bulls. Group 1, regarded as the normal class, had bulls within 60 days post-calving. In groups 2, 3 and 4, joining took place between 61-150, 151-230 and 231 or more days after calving respectively. In group 5, females did not conceive during the "first" breeding season but did so during the subsequent mating period. Preliminary analysis indicated that there was no additional information lost by excluding groups 3 and 5. Consequently, only groups 1, 2 and- 4 were retained for final evaluation.
Experiment 2: Effect of strategic supplementation on Calving Interval
Three experimental rations were prepared for supplementing in-calf cows during the dry period between January and March in 1978, 1979 and 1980 as follows:
|
Ration A: |
2 kg chopped fresh grass from the paddocks. |
|
Ration B: |
2 kg ground sorghum grain (Sorghum vulgare) mixed with 500 g of sugarcane molasses (Energy). |
|
Ration C: |
2 kg ground sorghum grain mixed with 500 g molasses and 75 g of urea (Energy + Nitrogen). |
The supplemented regimes were:
(i) chopped grass (control) - Ration A
(ii) pre-parturient supplementation(a) Energy alone - Ration B
(b) Energy + Nitrogen - Ration C(iii) Post-parturient supplementation - Ration 3.
Cows on feeding regime (i) and (ii) were supplemented during the last two months of pregnancy for varying periods until they calved down while those on regime (iii) were supplemented up to 60 days.
Experiment 3: Effect of Weaning on Calving Interval
Breeding females were suckling their calves each year. The calves were weaned in three batches at the end of September, October and November/December during 1977, 1978 and 1979 when they were between 5 and 9 1/2 months old.
Data
Each female was regarded as an entity during the breeding season and data were compiled to include breed, birth and calving dates, previous parous state i.e. whether the animal was a heifer, dry cow or suckling a calf at conception prior to the calving interval in question and conception period. The covariables included body weight at weaning (WWN), "earlier" calving (WCV), one month after calving (W1ACV), one and two months prior to "second" conception (W1BC and W2BC respectively) and at "second" conception (WCP), SMI1 and SMI2. From this information, age at "earlier" calving, calving interval and weight changes WWN-WCV, W1ACV-WCP, W1BC-W2BC and W1BC-WCP were computed. Factorial effects in experiments 1, 2 and 3 are summarised in Tables 1, 2 and 3 respectively.
Table 1. Data structure and number of females in experiment 1.
|
Class |
Levels |
Groups |
Total |
||
|
1 |
2 |
4 |
|||
|
1. Breed type |
Boran |
44 |
29 |
58 |
131 |
|
EASZ |
42 |
47 |
33 |
122 |
|
|
Crosses* |
28 |
20 |
7 |
55 |
|
|
2. Dam's age at earlier calving (years) |
3 or below |
18 |
16 |
22 |
56 |
|
4-8 |
65 |
38 |
71 |
174 |
|
|
9 and above |
31 |
42 |
5 |
78 |
|
|
3. "Second" conception period |
Mid Jan-mid March |
1 |
7 |
33 |
41 |
|
Mid March-mid May |
2 |
- |
8 |
10 |
|
|
Mid May-mid July |
52 |
59 |
19 |
130 |
|
|
Mid July-mid Sept |
37 |
26 |
1 |
64 |
|
|
Mid Sept-mid Nov |
6 |
- |
- |
6 |
|
|
Mid Nov-mid Jan |
16 |
4 |
37 |
57 |
|
|
4. Year of "second" conception |
1973 (early) |
2 |
11-70 |
83 |
|
|
1973 (late) |
26 |
- |
3 |
29 |
|
|
1974 |
2 |
3 |
- |
5 |
|
|
1975 |
28 |
59 |
19 |
106 |
|
|
1976 |
23 |
23 |
- |
46 |
|
|
1977 |
33 |
- |
6 |
39 |
|
|
5. Dam's previous parity |
Heifer |
17 |
17 |
31 |
65 |
|
Dry cow |
39 |
14 |
49 |
102 |
|
|
Suckled |
58 |
65 |
18 |
141 |
|
|
Total |
|
114 |
96 |
98 |
308 |
*Crosses were F1 from Boran and EASZ cows x Hereford bulls.
Table 2. Data structure and number of females in experiment 2.
|
Class |
Levels |
No. of observations |
|
1. Breed type |
Boran |
34 |
|
Boran x Hereford (F1) |
52 |
|
|
EASZ |
48 |
|
|
EASZ x Hereford (F1) |
51 |
|
|
2. Dam's age at earlier calving (yrs) |
4 |
22 |
|
5-9 |
100 |
|
|
10 and above |
63 |
|
|
3. Dam's previous parous state |
Dry cow |
72 |
|
Suckled |
113 |
|
|
4. Feeding regime |
Chopped fresh grass |
40 |
|
Pre-parturient supplementation |
57 |
|
|
(a) Energy |
43 |
|
|
(b) Energy + Nitrogen |
45 |
|
|
Post-parturient supplementation |
57 |
|
|
5. Month of earlier calving |
February |
83 |
|
March |
58 |
|
|
April/May |
44 |
|
|
6. Year of earlier calving |
1978 |
58 |
|
1979 |
53 |
|
|
1980 |
74 |
|
|
Total |
|
185 |
Table 3. Data structure and number-of females in experiment 3.
|
Class |
Levels |
No. of observations |
|
1. Breed type |
Boran |
21 |
|
Boran x Hereford (F1) |
42 |
|
|
EASZ |
29 |
|
|
EASZ x Hereford (F1) |
30 |
|
|
2. Dam's age at weaning (yrs) |
4 |
14 |
|
5-9 |
70 |
|
|
10 and above |
38 |
|
|
3. Weaning month |
End of September |
35 |
|
End of October |
38 |
|
|
End of Nov/December |
49 |
|
|
4. Weaning year |
1977 |
28 |
|
1978 |
43 |
|
|
1979 |
51 |
|
|
5. Calving month* |
Jan/February |
46 |
|
March |
36 |
|
|
April/May |
40 |
|
|
Total |
|
122 |
*Month of calving following weaning in question.
Statistical Analysis
The sources of variance and covariance for calving interval were analysed using the generalised least-squares regression methods for multiple classifications and non-orthogonal data as described by Harvey (1960) and adapted by Siebeck (1976) in his computer programme, SYSNOVA. All effects were considered fixed. The estimate of the mean squares attributable to any effect was computed after all other effects in the evaluation model had been fitted and was tested against that of the residual.
The generalised model used was:
Yijkl + ai + bj + cjk + (ab)ij + (ad)i + d(Dijkl-D) + eijkl
where,
Yijkl = calving interval of an individual cowYijkl = effect common to all cows
ai = effect of the ith A class after removal of
bj = effect of the jth B class after removal of
cjk = effect of the kth C class within the jth B class after removal of the jth B class. This was hierarchical effect of nutrients within the pre-parturient feeding regime.
(ab)ij = effect of the ijth AB sub-class after the average effects of A and B have been removed.
(ad)i = effect of an interaction between A and the continuous variate Dijkl. This interaction between a treatment and a covariate enables particular slopes to be fitted.
d = partial regression coefficient of calving interval Yijkl on Dijkl.
eijkl = random errors.
Weight Change During the Month Prior to "second" Conception
The effect of weight change on calving interval was influenced significantly (P<0.01) by the period during which females reconceived and is depicted in Figure 2. During the May-July period, animals reconceived at an average weight of 331 kg, and weight changes did not affect calving interval to any appreciable degree. Weight gains up to 10 kg prior to conception between July and September were associated with short calving intervals (Figure 2). Females reconceived between September and January at an average weight of 308 kg, and long intervals were associated with weight loss in excess of 10-15 kg. Influence of weight change also varied significantly (P<0.005) and curvilinearly depending on year of "second" conception as shown in Figure 3. While weight gain during 1976 resulted in systematic and sharp reduction in calving intervals, this effect was marginal during 1975. Weight gain during 1973/74 caused considerable variation in calving interval.
Weight Change Between One to Two Months Prior to "Second" Conception
The effect of this parameter was linear and highly seasonal (P<0.005) and is depicted in Figure 4. Females that gained weight prior to conception between July and September had calving intervals which were reduced by 0.7 days for each kg increase in body weight. This trend was reversed in the case of animals that conceived during periods other than July-September such that females that gained weight also had longer calving intervals. However, effect of weight gain prior to conception between May and July was marginal.
Figure 2. Relationship between calving interval and weight (W1BC-WCP) within conception period of group 1 cows.
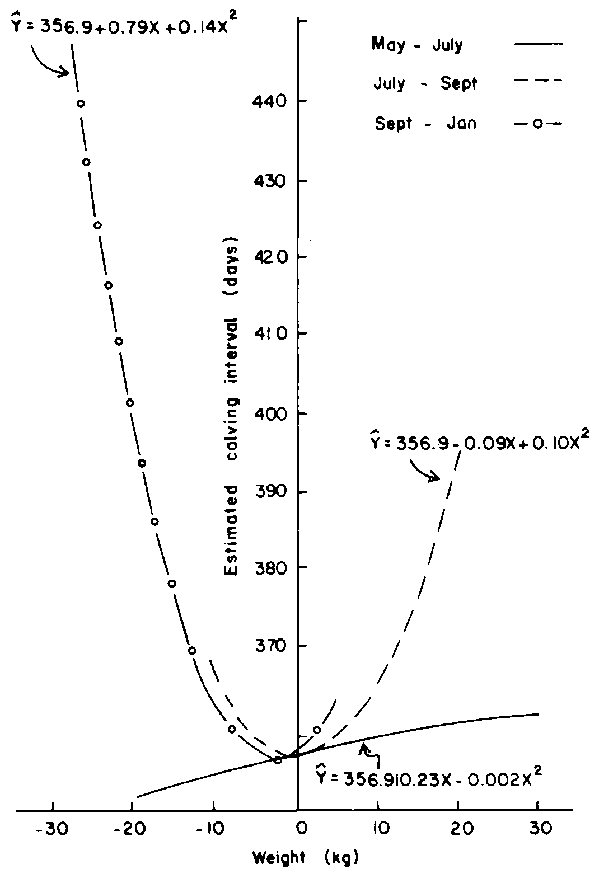
Figure 3. Relationship between calving interval and weight (W1BC-WCP) within conception year of group 2 cows.
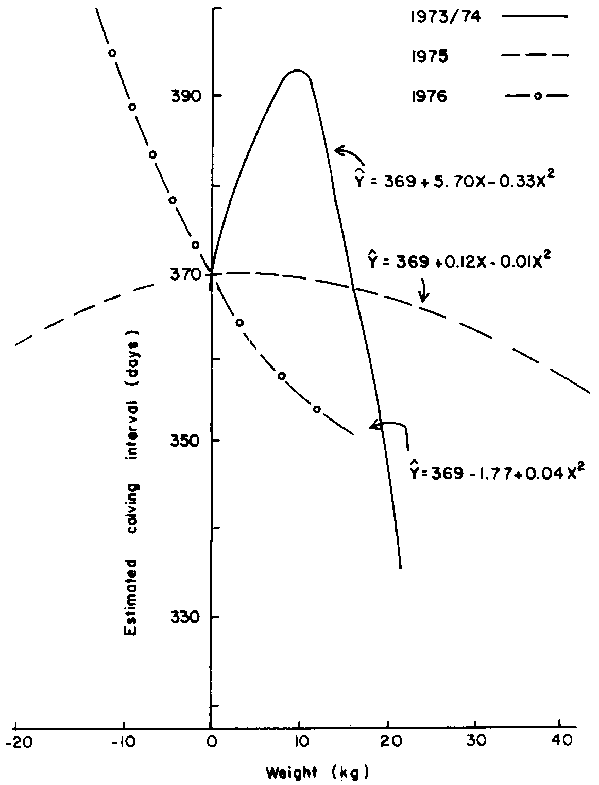
Figure 4. Relationship between calving interval and weight (W1CP-W2CP) within conception period of group 2 females.
Weight Change Between One Month After Calving and Reconception
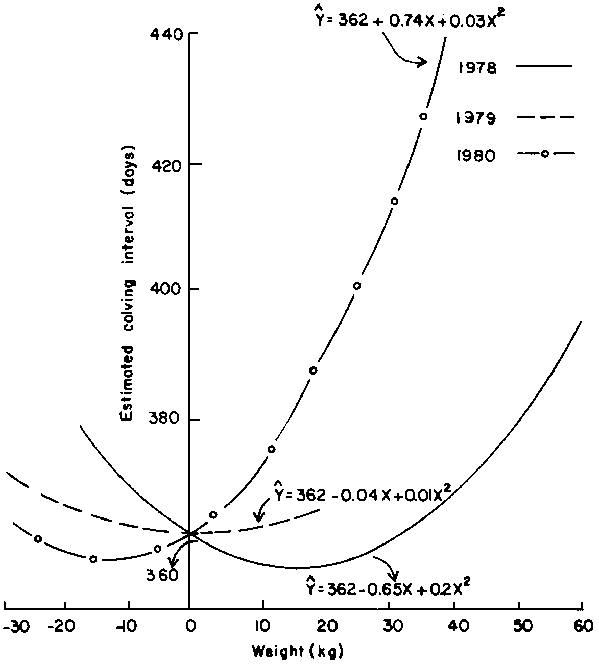
Influence of this factor varied significantly (P<0.05) depending on the year when reconception occurred as depicted in Figure 5. During 1978, weight gains up to 25 kg were associated with short calving intervals. During 1979, effect of weight change was marginal. Weight gains during 1980 resulted in long intervals. Mean calving interval was reduced by 0.7 days for each kg increase in body weight following calving during March; otherwise the overall effect of weight change did not vary across calving month.
Figure 5. Relationship between calving interval and weight (WIACV-WCP) within year of earlier calving of supplemental cattle.
Body Weight at "Second" Conception
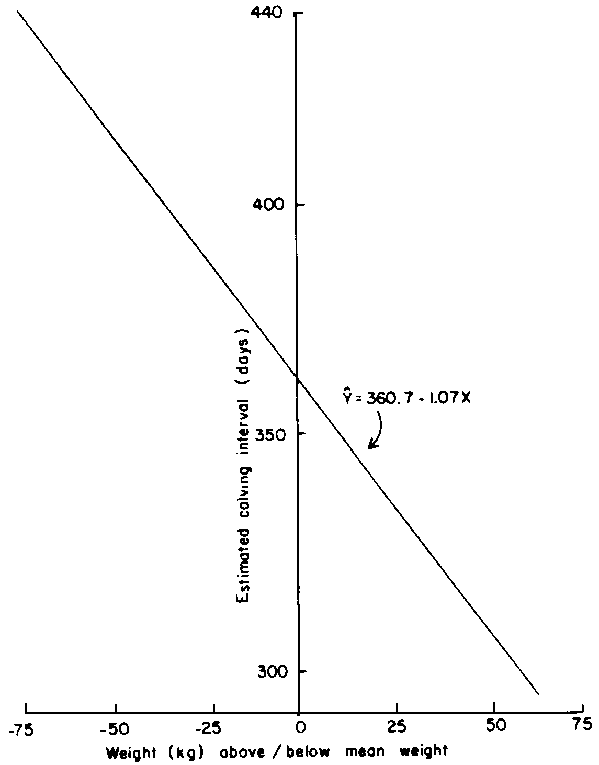
In the normal class of females, the optimal weight at "second" conception was 318 kg with a range between 310 and 330 kg (Figure 2). However, joining weight did not affect calving interval significantly. Group 4 females reconceived at an average weight of 311 kg, and heavier animals had a significantly (P<0.05) shorter mean calving interval. In the weaning trial, animals reconceived at an average weight of 351 kg, and for each kg increase in liveweight above the overall mean, there was a corresponding reduction (P<0.01) in calving interval of 1.1 days (Figure 6).
Figure 6. Relationship between calving interval and weight at "second" conception.
Soil Moisture Index
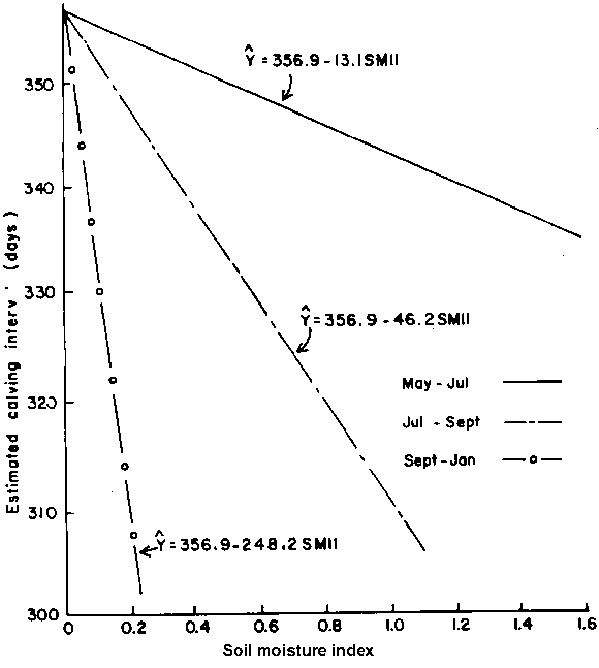
The effect of this factor on calving interval was linear and highly significant (P<0.005) but was modified by conception period as shown in Figure 7. In general, increase in SMI during the month prior to "second" conception was associated with a shorter mean calving interval. The response was greatest during the September-January conception period but was relatively marginal when conception occurred between May and September.
Figure 7. Relationship between calving interval and soil moisture index (SMII) within conception period of group 1 females.
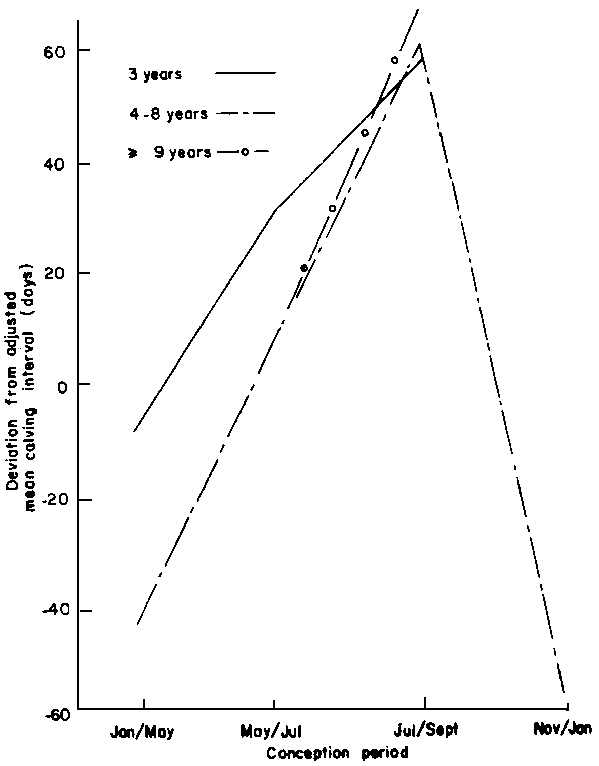
Age of Breeding Females
When evaluated within conception period, age of animals had a highly significant (P<0.005) influence on calving interval (Figure 8). The three-year-old females that conceived between January and July had a longer mean calving interval compared to the 4 to 8-year-old cows. The shortest intervals were achieved by the 4 to 8-year-old cows which conceived between November and January. Regardless of age, all animals that reconceived between July and September had long intervals.
Figure 8. Effect of age at calving (year) within conception period on calving interval of group 2 females.
Dam's Previous Parity
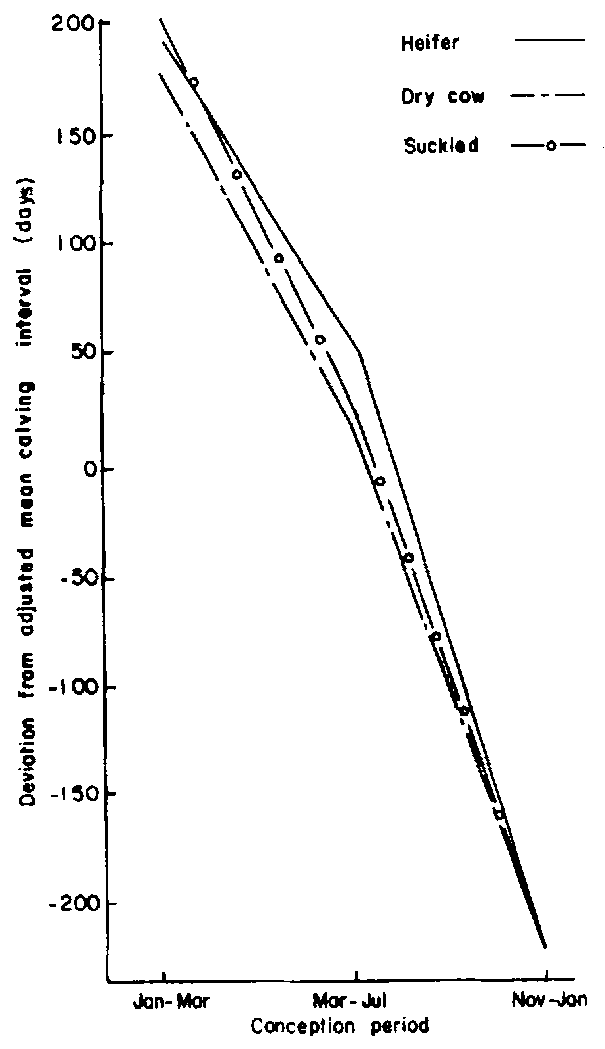
The effect of this parameter became marginally significant (P<0.05) when evaluated in the context of conception period (Figure 9). Heifers that conceived between January and July had longer calving intervals compared to previously dry cows. Reconception between November and January was associated with shorter calving intervals regardless of previous parity.
Year of "Second" Conception
Year effects on calving interval were highly significant (P<0.005). Females that conceived during 1973 (late), 1975 and 1977 had shorter intervals compared to those that conceived during 1973 (early) and 1976.
Role of Supplementation
Dry season supplementation had a significant effect on calving interval, but this was modified by a number of factors:
Age: Without supplementation, the 5 to 9-year-old cows had calving intervals which averaged 38 days shorter than the mean for the 4-year-old females (Figure 10). Supplementation, whether before or after calving, only maintained calving intervals of the mature cows at average levels. However, there was a dramatic reduction in the interval when the 4-year-old females were supplemented after calving.Previous parity: Females that suckled calves during the previous year responded positively to supplementation especially when given after calving (Figure 11).
Year of supplementation: While supplementation reduced calving intervals during 1978 and 1979 (post-parturient), it was virtually useless during 1980 (Figure 12).
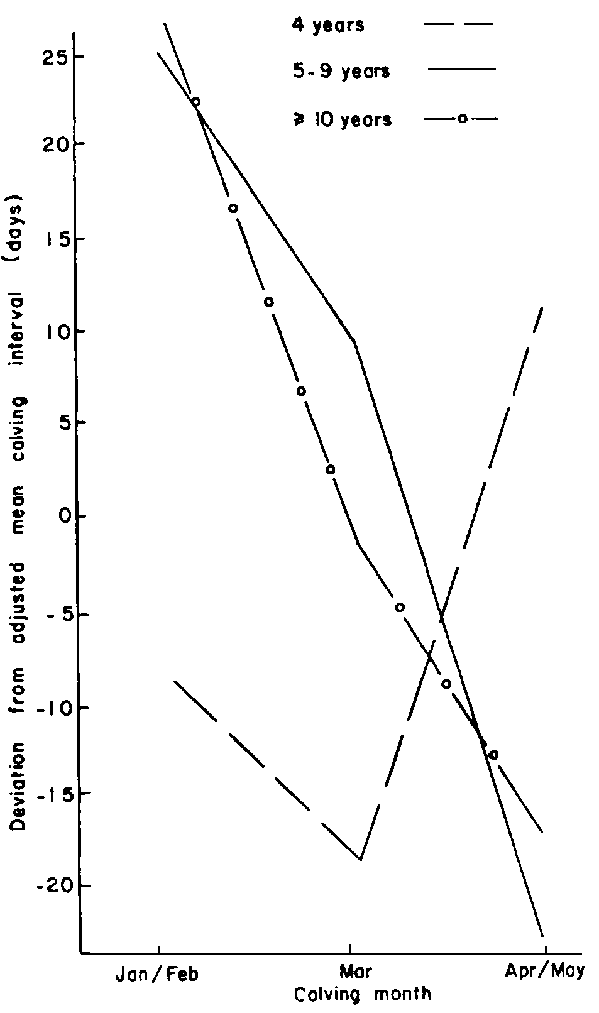
Month of "earlier" calving: Females that calved between March and early May had shorter calving intervals compared to those that calved during January and February (Figure 13). However, with no supplementation, the 4-year-old females that calved during April and May had long intervals (Figure 14).
Energy versus energy + urea: Differences in the effects of these nutrients when given pre-partum were not significant.
Figure 9. Effect of previous parity within conception period on calving interval of group 4 females.
Figure 10. Effect of age calving within feeding regime on calving interval of supplemental cattle.
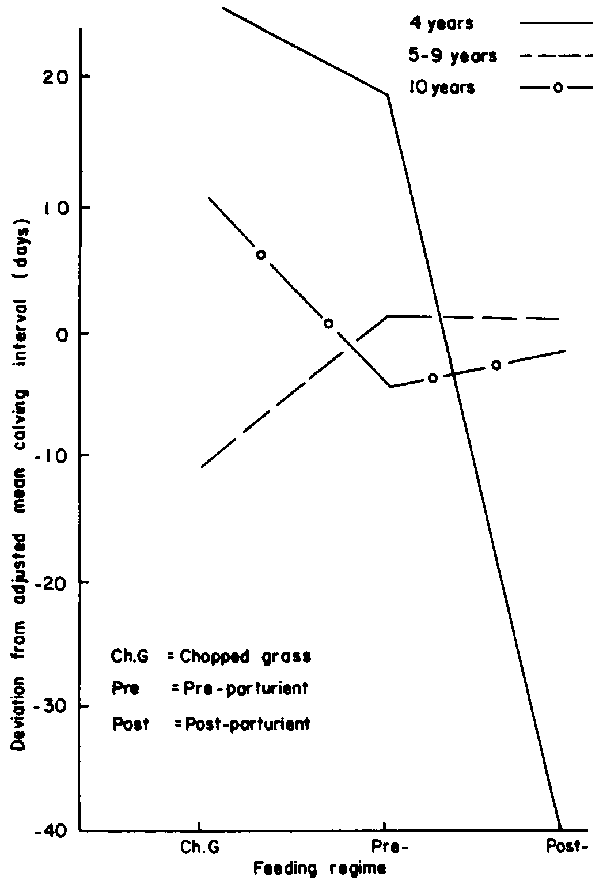
Figure 11. Effect of previous parity within feeding regime on calving interval.
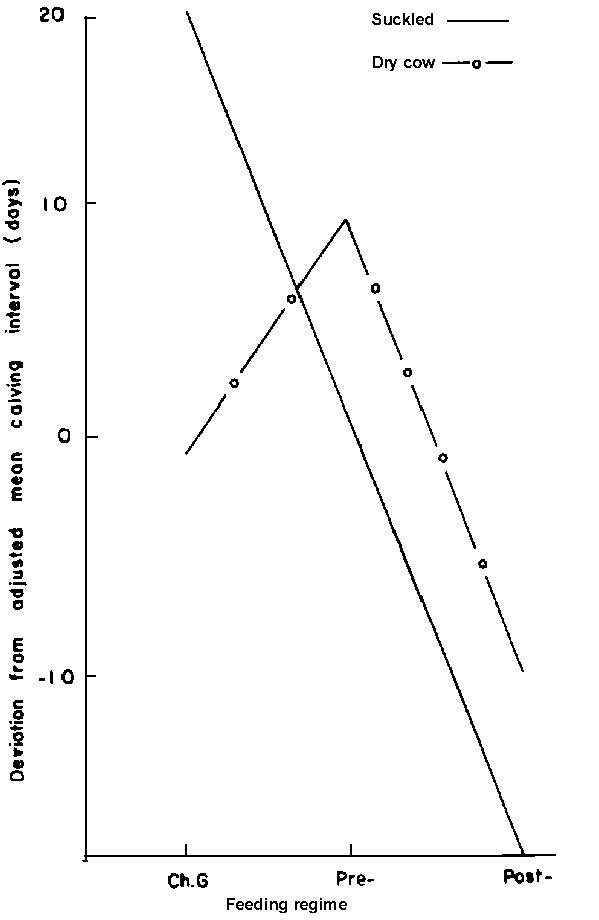
Figure 12. Effect of feeding regime within calving year on calving interval.
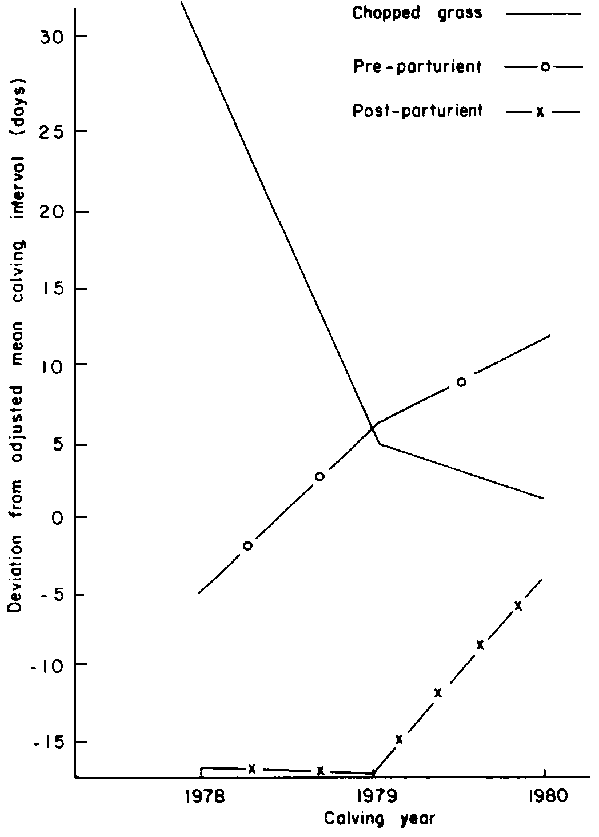
Figure 13. Effect of age (years) within month of earlier calving on calving interval of supplemented cattle.
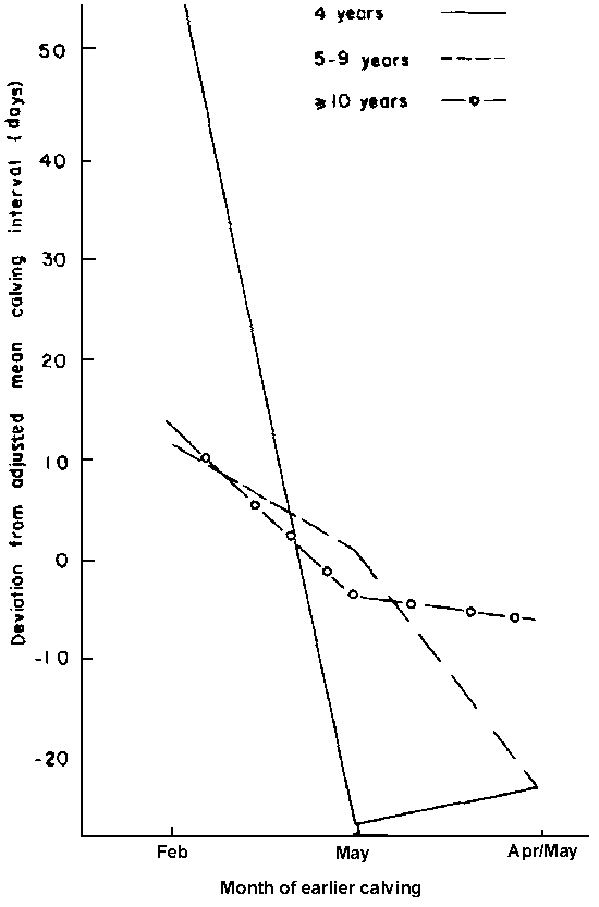
Figure 14. Effect of dam's age at weaning within subsequent calving month on calving interval.
Effects of Weaning
Calving intervals increased marginally (P<0.05) by 0.2 days for each day increase in the period before weaning was effected. There was some indication that the 4-year-old females that weaned calves at the end of September had a shorter mean calving interval than that of females that weaned calves at the end of October, otherwise the effect of weaning month per se was marginal (P<0.05). As weaning progressed from year to year, the 10-year-old-and-above cows had calving intervals which averaged 32 and 21 days longer than for the 4- and 5 to 9-year-old females respectively. In general, effects of weaning on calving interval were marginal.
Dam's Genotype
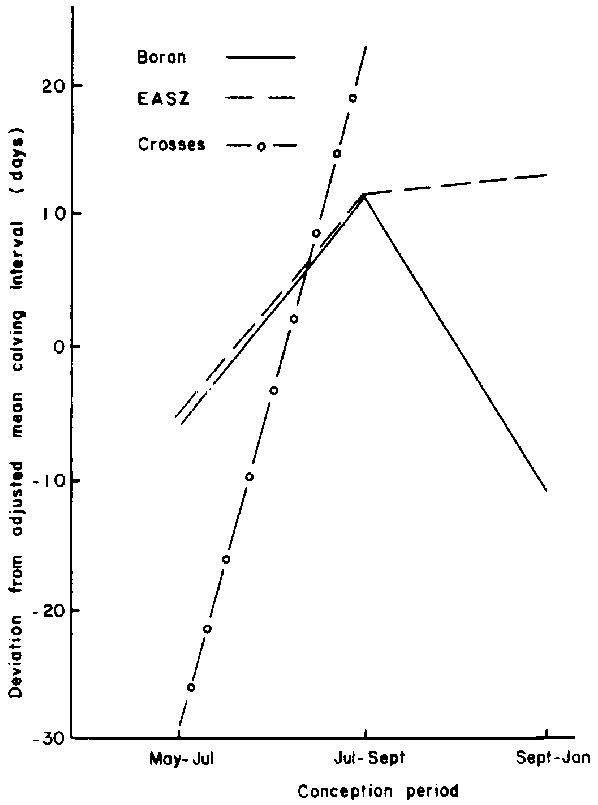
The influence of this parameter was significantly (P<0.05) modified by conception period (Figure 15). Both the EASZ and the Boran females had similar calving intervals except during the September-January conception period when the latter breed had shorter intervals. The crosses that conceived between May and July had calving intervals which averaged 30 days below the adjusted overall mean of 357 days. Animals of all genotypes which conceived between July and September had long intervals. With supplementation, the Boran crosses had significantly (P<0.01) shorter calving intervals than pure Borans.
Figure 15. Effect of breed type within conception period on calving interval of group 1 females.
The highly significant main effects and interactions with body weight changes, soil moisture index, age, previous parity and dam's genotype made conception period one of the most important factors affecting fertility of grazing beef cattle at Athi River Ranch. High fertility was achieved during the November-January conception period which normally covers the "short rains" with high temperatures. The high fertility observed between January and March (Figure 8) could be due to increased sexual activity mainly as a result of high temperatures (Anderson, 1944). However, in rather striking contrast, group 4 females that conceived during the same period had a long fertility (Figure 9). Considering that the number of females that conceived during this period in group 4 was much greater than that in group 2 (Table 1), it would appear that inadequate grazing had a more marked adverse effect on fertility than the influence of increased temperature and sexual activity. Low fertility observed during the cold and dry period between July and September could be due to a level of intake sufficient to bring about ovulation but grossly inadequate for full manifestation of oestrus.
The highly significant year effects reflected important variation in fertility from year to year. Admittedly, a large proportion of year effects would be confounded with variation associated with conception period. But since it is possible that factors other than conception period could contribute significantly to the observed variation, inclusion of year effects in the evaluation model would still be justified. Age of bulls and management practices relating to stocking rate, mineral supplementation and culling programmes are some of the factors which could vary from year to year.
Bishop (1978) reported high conception rates following rainfall during the previous year. However, in Kenyan rangelands with such tremendous variability in rainfall both annually and seasonally (French, 1957), it would be desirable to correlate fertility with rainfall within the immediate past so that corrective measures could be instituted in good time. In Australia, Andrews (1976) reported a high correlation between conception rate and rainfall with a one-month lag. Working at Athi River Ranch, Potter (1985) observed that liveweight gain was correlated with rainfall occurring between three to six weeks previously. The period of SMI evaluation in this study coincided with the one used by Andrews (1976) but did not quite correspond with the one evaluated by Potter (1985). Further work is needed to pin-point this period more precisely. Potter (1985) has suggested that use of rainfall data was just as good as the more elaborate and often cumbersome water balance information involving evapotranspiration data. Although rainfall may be an important factor, Andrews (1976) contended that a period of one month was not sufficient time for pastures to grow and improve body condition and hence conception rate. He postulated that increased fertility was likely to be due to either a change in the nutritional composition of the pastures or an increase in the rate of intake or both through the action of the hypothalmus and/or the pituitary. However, it is important to note that rangeland vegetation at Athi River Ranch grows very fast and matures very quickly (Karue, 1972; 1974), and so it may have a short-term direct effect on body condition and hence fertility.
The debate continues as to whether the animal's ability to conceive was a function of its absolute body weight per se (Lammond, 1970; Grosskopf, 1980) or weight change prior to conception (Elliot, 1964, cited by Topps, 1977; Capper et al., 1977; Thorpe et al., 1981). While suggesting the concept of a target joining weight, Lammond (1970) had intimated that for each cow, depending on breed, age and year, there was a certain range in body weight and body condition required for conception. By implication, therefore, it did not matter whether animals lost or gained weight during the breeding season provided their liveweight was maintained above a certain minimum. The ability to maintain weight would depend, among other things, on the current lactational stress on the animal, hence the reason why researchers evaluate body weight in heifers, dry or lactating cows (Kidner, 1966; Sacker et al., 1971; Morley et al, 1976). Such evaluation would be possible if fertility was considered in terms of calving date rather than calving interval. Another aspect is whether the various workers were evaluating absolute or proportionate weight changes. A proportionate weight change such as that used in this study with weight at "second" conception as a reference point would render changes in small animals (though of the same breed) comparable to similar changes in bigger ones. It seems as if both body weight and weight change operate singly or simultaneously to influence fertility depending on the lactational and/or nutritional status of the animal. For instance, although animals in the normal class reconceived at an average weight of 320 kg well within the optimum range of 310 to 330 kg for this group of animals, influence of body weight was not significant. This could have been due to the adverse effect of suckling reported by Wiltbank and Cook (1958) and Hutchison (1983).
Group 4 females had dried off at the time they reconceived, and the effect of joining weight was able to be manifested, though marginally due to the fact that animals conceived at an average weight of 311 kg which was at the bottom of the required range in body weight. Under these circumstances, both body weight at joining and weight change affected fertility simultaneously. Animals reconceived between September and January and during 1976 at average weights of 308 and 298 kg respectively. Both weights were below the minimum weight required for conception, and animals that had a higher fertility were those that gained or lost less body weight during the month prior to conception. With favourable nutritional conditions between May and July (Figure 2), during 1975 (Figure 3) and 1979 (Figure 5), animals reconceived at average weights of 330, 311 (half the number of females were the small EASZ) and 352 kg respectively. Under these conditions, effect of weight change was marginal and animals could gain or lose weight without jeopardising their fertility. The picture during 1973/74 (Figure 3) was somewhat confusing probably due to the fact that fertility was evaluated over a two-year period. In any case, the sample size during 1973 and 1974 was far too small to warrant any serious consideration of the effects during these two years. Curvilinearity in response of weight change could also be ignored.
Dry cows (group 4) and females that had settled down after calving before they were re-introduced to the bulls (group 2) tended to gain weight prior to conception to the detriment of fertility. However, in normal ranching situations, animals would be lactating at the time they reconceive with minimum chances of overweight. Failure to register significant effects of weight change between one month after calving and conception could have been due to the rather limited variation in climatic conditions during the pre-selected breeding season between May and July. However, it was evident that weight changes occurring near conception exerted a much greater effect on fertility than changes far removed from conception. What is, perhaps, perplexing is the revelation by Thorpe et al. (1981) that liveweight was merely an approximate indicator of the nutritional status of a beef animal. These workers postulated the existence of an intricate relationship between nutrition, hormonal control, lactation and fertility in which the post-partum period with its associated lactational stress was considered more important than variation due to liveweight. Since this relationship is still vaguely understood, monitoring of body condition as suggested by Andrews (1976) together with proportionate body weight changes during the month prior to breeding would give a reasonable indication of the fertility status of a grazing beef animal.
Supplementation of beef cattle with energy (Wiltbank et al 1962 and 1964; Dunn et al., 1969) or nitrogen (Elliot, cited by Christie, 1962; Siebert et al., 1976; Holroyd et al., 1977) has been a normal practice to improve fertility but results have not been consistent. For instance, in Botswana, Capper et al. (1977) observed no response to stimulatory licks of molasses and urea by dry cows or cows with calves older than five months. More recently, Holroyd et al. (1983) supplemented beef cows with urea and molasses but observed no effect on fertility. In this study, improved fertility was observed in the four-year-old and previously suckled females. It seems imperative, therefore, that assessment of the effect of supplementation should be done in the context of age and previous parity. Failure to detect differences in the effect of grain versus grain plus urea feeding could be attributed to the favourable climatic conditions especially during 1979 and 1980 which made it possible for animals to derive most of their nutrient requirements from pasture alone. It is worth noting that the last trimester of pregnancies at the ranch coincided with the period covering the "short rains" between November and December. Under such conditions, Orskov (1982) suggested that nitrogen requirements for pregnancy could be met by microbial protein synthesis alone. However, there is a tremendous increase in the nutritional requirements for lactation and reconception (Crampton and Lloyd, 1959), hence the significance of post-calving supplementation. It would, therefore, be of interest to investigate the effect of nitrogen and energy sources when given post-partum. Since all indications pointed at conception as the most critical period, it would be necessary to pinpoint more precisely how soon after calving should supplementation be given. Because of the intricate relationship between non-protein nitrogen (NPN) and carbohydrate metabolism and utilisation (Orskov, 1982), further investigations should be focused not on the relative importance of NPN and energy but rather on the proportion of rumen-degradable and undegradable nitrogen relative to metabolisable energy intake.
The effect of age on fertility of beef cattle is an indirect one through its influence on body condition to which young cattle are more sensitive (Andrews, 1976). As body condition was related to liveweight, Wiltbank and Spitzer (1978) have stressed the significance of allowing young cattle to attain a target weight and age before first mating - the weight and age varying according to breed. Under East African conditions, Macfarlane and Worrall (1970) had indicated that puberty in Boran heifers occurred at approximately 60% of the mature body weight. However, mere attainment of puberty is not enough as young cattle are expected to conceive, carry the foetus to full term, rear the calf successfully and reconceive at the earliest opportunity. It is possible that the 250 kg liveweight stipulated for first breeding of heifers at the ranch was lower than the threshold joining weight especially in crosses. This would not only delay reconception but could curtail their productive life. If anything, it would be better to breed heifers too late than too soon. Young beef cattle were more vulnerable to adverse nutritional conditions at post-partum oestrus. However, their fertility improved as the post-calving period before joining with the bulls increased from about five months onwards. There was evidence from the weaning trial that highest fertility was achieved by the four-year-old females. The implication of these observations is that young cattle should be allowed about five months after calving before joining with bulls for their second mating season. This would mean that first breeding in heifers and that of the main herd would be out-of-phase, although all indications were in favour of seasonal mating. However, earlier breeding in heifers would necessitate their separation from the main herd which would facilitate closer attention and feeding to ensure high fertility. A suggested package for young beef cattle would be (a) mating between November and January or, in general, immediately after a rainy period (b) separating and mating approximately three months before the main herd to synchronize second mating with the rest of the herd and (c) providing a supplementary diet after first calving.
The timing of the breeding programme at the ranch did not permit evaluation of the effects of calving months other than those between late January and early May. However, it was evident that animals that calved down during the dry season in January and February had a low fertility. This could be due to failure to regain lost body weight (Kidner, 1966) but as Christie (1962) had indicated, dry season loss in body weight per se did not, necessarily, reduce fertility as long as the animal was able to recoup during the subsequent wet season. Animals that calved down during March had a high fertility which confirmed Daly's (1971) suggestion that calves should be dropped at the end of a dry season. The older cattle that calved down during the rainy season in April-May seemed to get away with less favourable nutritional conditions at post-partum oestrus probably because of their ability to draw on body reserves accumulated during the wet period (Orskov, 1982).
There were indications to suggest that previous parity had an influence, albeit small, on fertility. Results from various workers reporting on this parameter are rather inconsistent (Christie, 1962; Buck et al., 1976; Thorpe et al., 1981). It would appear that in order to make meaningful comparison in fertility, one would have to consider previous parity in the context of conception period. When nutrition was adequate, for instance during November-January conception period, or when supplementation was given after calving, influence of previous parity became insignificant. Generally, carry-over effects arising from previous parity were of less importance in the fertility complex. This should not be confused with the overwhelming influence from current lactational stress on the animal (Crampton and Lloyd, 1959; Trail, 1968), and the whole idea behind weaning is to remove this stress thereby ensuring that animals attain the body weight and body condition required for reconception. Lammond (1970) had suggested increasing body weight and condition before calving and attempting to hold this condition up to the breeding season. This could be achieved by early weaning but because of seasonality in pasture productivity (Karue, 1972, 1974), it would be difficult to maintain body weight. Besides, results of this study indicated that fertility was not always correlated with body weight at calving. The other alternative also suggested by Lammond (1970) was to allow cows to calve down in reasonable body condition and feed heavily up to conception. This would be a much more practical method which was supported by results of this study relating to post-partum supplementation.
The question of weaning month necessitated some consideration because of the observed sensitity to pasture by the early weaned calves (Schotter and Williams, 1975) on one hand and the need to prepare the breeding female for subsequent calving and, more importantly, reconception on the other. However, effects of weaning month were, at best, only marginal probably because of a closed breeding period and hence weaning season. Thus, in considering the effects of lactation, it would be more important to pay particular attention to when animals are due to calve down so that they can be supplemented for early reconception rather than attempting to increase body weight and body condition at calving by early weaning.
The EASZ females exhibited fairly consistent fertility levels throughout the year, an observation that was consistent with the findings of Galukande et al. (1962). Zebu cattle are indigenous in East Africa (Mason and Maule, 1960) and would be expected to have adapted themselves to fluctuations in feed supply and related factors from out-of-season rains and prolonged droughts. Adaptability has been attributed to their ability to lay down fat on a lower plane of nutrition (Mason and Buvanendran, 1982). Fertility of the Borans was, by and large, similar to that of the EASZ which supported the observations reported by Trail (1968) at Ruhengere Ranch in Uganda. However, there was evidence to indicate that the Borans had a higher fertility during the last quarter of the year when pasture productivity was adequate.
Results of cross-breeding to improve fertility have been inconsistent. Some reports have indicated a reduction in age at first calving (Mahadevan and Hutchison, 1964) and increased manifestation of oestrus in cross-bred zebu cattle (Swensson et al., 1981). Observations from this study tended to tip the balance in favour of Boran crosses under conditions of adequate nutrition. High fertility could be due to heterosis. However, this could not be confirmed due to lack of reciprocal crosses. Although it is desirable to maintain the indigenous zebu germplasm, the prospects of increased beef productivity from rangelands based on the Boran and its crosses look very promising.
Fertility in range-fed beef cattle is a very complex physiological process involving the interplay of a number of factors. These factors are overwhelmingly climatic in origin which means that environmental conditions especially nutrition and management have to be tailored for the benefit of the animal. Nutritional and management factors immediately before and during the mating season had the most significant and far-reaching effects on fertility. Even during the conception period, influence of nutrition and related factors such as rainfall, body weight and dam's age varied from season to season and was modified by other factors which, all working in concert, complicated the situation even further.
Fertility was seasonal. Consequently, seasonal mating would be advocated. High fertility was observed between November and January followed by May-July. The significant year effects reflected mainly seasonal effects although variation across years could be attributed to changes in feeding, mating practices and age of bulls. Increased rainfall during the month before conception was associated with high fertility. The effect was seasonal which underscored the significance of conception period and seasonal mating.
The target joining weight was around 318 kg with a range between 310 and 330 kg. Heavier cows at conception were more fertile than lighter ones, but the depressing effect of suckling could counteract that of body weight unless joining weight was well above 318 kg. Animals that gained body weight or suffered less weight loss during the month prior to conception had a higher fertility. Previously dry cows tended to gain weight prior to conception and gave a better indication of fertility status than weight changes far-removed from conception.
Dry season supplementation only helped to maintain fertility of older cows at average levels. However, it improved fertility of young and previously suckled females especially when given post-partum. There was no significant difference in the fertility of cows that were supplemented before calving with energy or energy plus urea.
Fertility improved marginally following early weaning. Some benefit was achieved when young females weaned their calves in September following a dry and cold season. Generally, effects of weaning were of less significance in the fertility complex.
Fertility also improved as the dam's age increased from three years onwards but declined after about nine to ten years. Young cattle were more vulnerable to adverse post-partum nutritional conditions. However, their fertility improved when they were allowed about five months after first calving before mating for the second parity. This would mean separation and mating of heifers some three months or so before breeding in the main herd to be able to synchronise their second mating with the rest of the herd.
Effect of previous parity on fertility was marginal depending on the nutritional level during the breeding season. Under grazing conditions without supplementation, dry cows were more fertile than previously suckled animals. This trend was reversed with supplementation. Females that conceived between November and January had a high fertility regardless of previous parity. Fertility levels of the East African shorthorn Zebu and Boran cows were similar except during the last quarter of the year when the latter breed had a better performance. Generally, crosses had a high fertility following a rainy season. However, with dry season supplementation, performance of the Boran crosses excelled the rest of the genotypes.
This paper was prepared for presentation at the PANESA 1987 workshop with the permission of the Director, K.A.R.I.
Anderson, J. 1944. The periodicity and duration of oestrus inb Zebu and grade cattle. Journal of Agricultural Science (Cambridge) 34:57-68.
Andrews, L.G. 1976. Reproductive performance of beef cattle in the Northern Territory. M.Sc. thesis, James Cook University, Australia.
Bishop, E.J.B. 1978. The performance of a beef breeding herd subjected to continuous mating in the Valley Bushveld of the Eastern Cape. South African Journal of Animal Science 8:15-25.
Buck, N.G., Light, D., Rutherford, A., Miller, M., Rennie, T.W., Pratchett, D., Capper, B.S. and Trail, J.C.M. 1976. Environmental factors affecting beef cow reproductive performance in Botswana. Animal Production 23:357-363.
Capper, B.S., Pratchett, D., Rennie, T.W., Light, D., Rutherford, A., Miller, M., Buck, N.G. and Trail, J.C.M. 1977. Effect of rumen stimulating licks on reproductive performance and liveweight gain of beef cattle in Botswana. Animal Production 24:49-55.
Christie, G.J. 1962. Sterility or reduced fertility due to nutritional causes. Bulletin of Epizootic Diseases of Africa 10:161-167.
Crampton, E.W. and Lloyd, L.E. 1959. General nutritional aspects of reproduction and lactation. In: Fundamentals of nutrition. W.H. Freeman and Co., San Francisco and London. pp. 386-393.
Daly, J.J. 1971. A breeding policy for beef herds. Queensland Agricultural Journal 97: 171-175; 237-242.
Dunn, T.G., Ingalls, J.E., Zimmerman, D.R. and Wiltbank, J.N. 1969. Reproductive performance of two-year-old Hereford and Angus heifers as influenced by pre- and post-calving energy intake. Journal of Animal Science 29:719-726.
French, M.H. 1957. Nutritional value of tropical grasses and fodders. Herbage Abstracts 27:1-9.
Galukande, E.B., Mahadevan, P. and Black, J.G. 1962. Milk production in East African Zebu cattle. Animal Production 4:329-336.
Gethin-Jones, G.H. and Scott, R.M. 1955. A second provisional soil map of Kenya and Tanganyika. East African Agriculture and Forestry Research Organization, Muguga, Kenya.
Grosskopf, J.F. 1980. Non-pathogenic factors associated with the reconception of beef cows under extensive conditions. In: IV World Conference on Animal Production. The Argentina Association of Animal Production, Buenos Aires. pp. 511-525.
Harvey, W.R. 1960. Least squares analysis of data with unequal subclass numbers. Agricultural Research Service, U.S.D.A., Wash., D.C.
Holroyd, R.G., Allan, P.J. and O'Rourke, P.K. 1977. Effect of pasture type and supplementary feeding on the reproductive performance of cattle in the dry tropics of North Queensland. Australian Journal of Experimental Agriculture and Animal Husbandry 17:197-206.
Holroyd, R.G., O'Rourke, P.K., Clarke, M.R. and Loxton, I.D. 1983. Influence of pasture type and supplement on fertility and liveweight of cows and progeny growth rate in the dry tropics of Northern Queensland. Australian Journal of Experimental Agriculture and Animal Husbandry 23:4-13.
Hutchison, H.G. 1983. Physiological limitations on reproduction performance: In: World Conference on Animal Production. European Association for Animal Production, Rome. pp. 147-148
Hutchison, H.G. and Macfarlane, J.S. 1958. Variation in gestation periods of Zebu cattle under ranch conditions. East African Agricultural and Forestry Journal 24:148-152.
Karue, C.N. 1972. Supplementary feeding of beef cattle. Paper presented to the Animal Production Society of Kenya, October.
Karue, C.N. 1974. The nutritive value of herbage in semi-arid lands of East Africa. 1. Chemical composition. East African Agricultural and Forestry Journal 40:89-95.
Kidner, E.M. 1966. Seasonal body weight changes in suckled Boran cows at pasture and their probable association with interval between carvings. East African Agricultural and Forestry Journal 31:399-404.
Lammond, D.R. 1970. The influence of undernutrition on production in the cow. Animal Breeding Abstracts 38:359-372.
Ledger, H.P., Odero, J.C. and Ndugire, N. 1969. Athi River Field Station. Record of Research Annual Report. East African Agriculture and Forestry Research Organization, Muguga, Kenya. pp. 118-119.
Livestock Development Division. 1983. Annual Report. Ministry of Agriculture and Livestock Development, Kenya. pp. 25-27.
Macfarlane, J.S. and Worrall, K. 1970. Observations on the occurrence of puberty in Bos indicus heifers. East African Agricultural and Forestry Journal 15:409-410.
Mahadevan, P. and Hutchison, H.G. 1964. The performance of crosses of Bos taurus cattle for milk production in the coastal region of Tanganyika. Animal Production 6:331-336.
Mason, I.L. and Maule, J.P. 1960. The indigenous livestock of Eastern and Southern Africa. Commonwealth Agricultural Bureaux, Farnham Royal, Buckinghamshire.
Mason, I.L. and Buvanendran, V. 1982. Breeding plans for ruminant livestock in the tropics. FAO, Rome.
McCulloch, J.S.G. 1965. Tables for the rapid computation of the Penman estimate of evaporation. East African Agricultural and Forestry Journal 30:286-295.
Morley, F.H.W., Axelsen, A. and Cunningham, R.B. 1976. Livestock at joining and fertility in beef cattle. Proceedings of the Australian Society of Animal Production 11:201-204.
Orskov, E.R. 1982. Host animal protein requirements and protein utilization. In: Protein nutrition in ruminants. Academic Press Inc. (London) Ltd., London. pp. 85-136 151.
Pereira, H.C. and McCulloch J.S.G. 1962. Water requirements of East African crops. In: E.W. Russell (ed.), The national resources of East Africa. D.A. Hawkins Ltd., Nairobi. pp. 88-91.
Potter, H.L. 1985. Aspects of climate, herbage growth and animal production in a semi-arid area of Kenya. Ph.D. thesis, University of Nairobi.
Sacker, G.D., Trail, J.C.M. and Fisher, I.L. 1971. Crossbreeding beef cattle in Western Uganda. 4. Calving percentage and mothering ability. Animal Production 13:165-170.
Schotter, J.H. and Williams, W.J. 1975. The effects of early weaning of Brahman-cross calves on calf growth and reproductive performance of the dam. Australian Journal of Experimental Agriculture and Animal Husbandry 15:456-459.
Siebeck, R.M. 1976. SYSNOVA version: a reference manual. Tropical Research Centre, Division of Animal Production, C.S.I.R.O. Rockhampton, Queensland.
Siebert, B.D., Playne, M.J. and Edye, L.A. 1976. The effects of climate and nutrient supplementation on the fertility of heifers in North Queensland. Proceedings of the Australian Society of Animal Production 11:249-253.
Swensson, C., Schaar, J., Brannang, E. and Meskell, L.B. 1981. Breeding activities of the Ethio-Swedish integrated rural development project. III. Reproduction performance of Zebu and crossbred cattle. World Animal Review (F.A.O.) 38:31-36.
Thorpe, W., Cruickshank, D.K.R. and Thompson, R. 1981. Genetic and environmental influences on beef cattle production in Zambia. Animal Production 33:165-177.
Topps, J.H. 1977. The relationship between reproduction and undernutrition in beef cattle. World Review of Animal Production 13:43-49.
Trail, J.C.M. 1968. Beef breeding at the Ruhengere Field Station. Reproductive performance. In: G.D. Sacker and J.C.M. Trail (eds). Proceedings of the beef cattle breeding and ranching development conference, Mbarara. Ministry of Animal Industry, Game and Fisheries, Uganda. pp. 44-49.
Wiltbank, J.N. and Cook, A.C. 1958. The comparative reproductive performance of nursed cows and milked cows. Journal of Animal Science 17:640-648.
Wiltbank, J.N., Rowden, W.W., Ingalls, J.E., Gregory, K.E. and Koch, R.M. 1962. Effect of energy level on reproductive phenomena of mature Hereford cows. Journal of Animal Science 21:219-225.
Wiltbank, J.N., Rowden, W.W., Ingalls, J.E. and Zimmerman, D.R. 1964. Influence of post-partum energy level on reproductive performance of Hereford cows restricted in energy intake prior to calving. Journal of Animal Science 23:1049-1053.
Wiltbank, J.N. and Spitzer, J.C. 1978. Recent research on controlled reproduction in beef cattle. Practical application. World Animal Review (F.A.O.) 27:30-35.
![]()
![]()
![]()
{kind=link}
{kind=link}