![]()
![]()
![]()
E.K. Maranga
National Range Research Station, Kiboko
P.O. Box 12, Makindu, Kenya
Introduction
Materials and methods
Results
Discussion
Conclusions
Acknowledgements
References
Abstract
Seasonal patterns of transpiration, leaf conductance and leaf water potential of herbaceous and shrub plant species in semiarid savanna ecosystems were determined over a growing season. Transpirational use of water for the shrub species ranged between 20 micrograms/cm-2 S-1 and 30 micrograms/cm-2 S-1. The deep-rooted shrub species maintained favourable leaf water balance and remained green over the entire growing season. The water response curves for the shallow-rooted plants indicated that they had little control over their water economy. Environmental processes leading to bush encroachment seem to be closely related to water response patterns of semi-arid savanna ecosystems in Kenya's pastoral grazing lands.
Savanna grasslands in Kenya occupy areas of marginal rainfall that constitute over one half of the total area of the country (Pratt et al., 1966). Water availability is the major driving variable controlling biological activity and regulating productivity of savanna ecosystems (Walker et al., 1981; Maranga et al., 1983; Maranga, 1986). Rainfall events are sporadic and often occur in irregular pulsations of short-lived storms (Griffiths, 1972). Effective rainfall recharges soil moisture for a specific period of time. The time elapsing between a change in soil-water status and an observed change in a plant process constitutes the response period (Sala et al., 1982). The response period of water-controlled processes is not fixed but is, at least partially, a function of the degree of water stress to which plants were previously exposed. The efficiency of utilisation of water input will decrease as the response period increases and therefore will depend on the water status (leaf water potential) of the plants prior to a rainfall event (Sala et al., 1982).
This paper attempts to evaluate water responses of native shrub and grass species to fluctuating soil water in the semiarid lands of Kenya. Plant variables used in this study to gauge plant water responses include leaf water potential and transpiration both of which are important determinants of plant productivity in semi-arid areas. The adaptive mechanisms that explain the observed responses will be deciphered.
Data and conclusions presented here come from field experiments conducted at the National Range Research Station, Kiboko. Mean annual rainfall is 600 mm based on over 70 years of data from Makindu Meteorological Station located near the research station. Rainfall is bimodally distributed. The "long" rains last from March to May; the "short" rains begin in or around October and end in December.
Detailed descriptions of the vegetation physiognomy of the study area are documented by Michieka and van der Pouw (1977). The vegetation of the experimental area can be described generally as open-wooded grasslands. There are scattered trees of Acacia seyal, Acacia tortilis and Commiphora riparia. There is a patchy understorey of shrubs of Grewia sp., Duosperma sp. and other genera scattered in a matrix of grassland characterised by Themeda triandra, Panicum maximum, Digitaria macroblephara and Chloris roxburghiana.
Field studies of soil water potential distribution with depth on sites that were dominated by Digitaria macroblephara, Panicum maximum, Acacia tortilis, Commiphora riparia, Grewia villosa and Hermania alhensis were carried out by means of a modified version of Spanner's (Spanner, 1951) Peltier thermocouple psychrometer installed at 10, 30, 60, 90 and 120 cm depths. The Peltier psychrometer system consisted of a microvoltmeter system, a control unit to supply current for Peltier cooling and a thermo couple psychrometer. The Hewlett Packard Model 419A (Hewlett Packard Englewood, Colorado) dew point microvoltmeter was used in conjunction with the thermocouple psychrometers calibrated for the five depths. Transpiration and diffusive resistance were measured by means of a steady state porometer (Li-Cor LI-1600 diffusive resistance porometer) between 10.00 a.m. and 2.00 p.m.
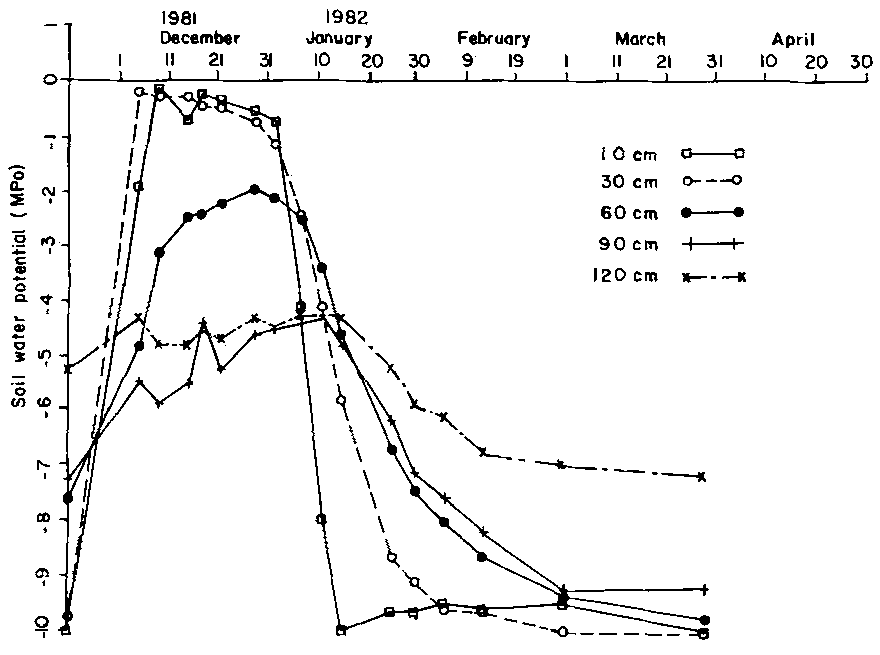
Effective rainfall contribution recharged the dry soils increasing soil water potentials from -10 MPa in early December to field capacity conditions by mid-December in the surface depths of 10 cm and 30 cm. Soil water potential in the deeper depths of 60-120 cm oscillated between approximately -7 MPa and 3 MPa over the same period (Figure 1). The wetting phase in the short rainfall season maintained field capacity conditions in the surface depths for a period of approximately three weeks. As soil water input declined, a sharp drop in soil moisture potentials occurred for the shallower soil profiles with only a gradual decline for the deeper soil horizons.
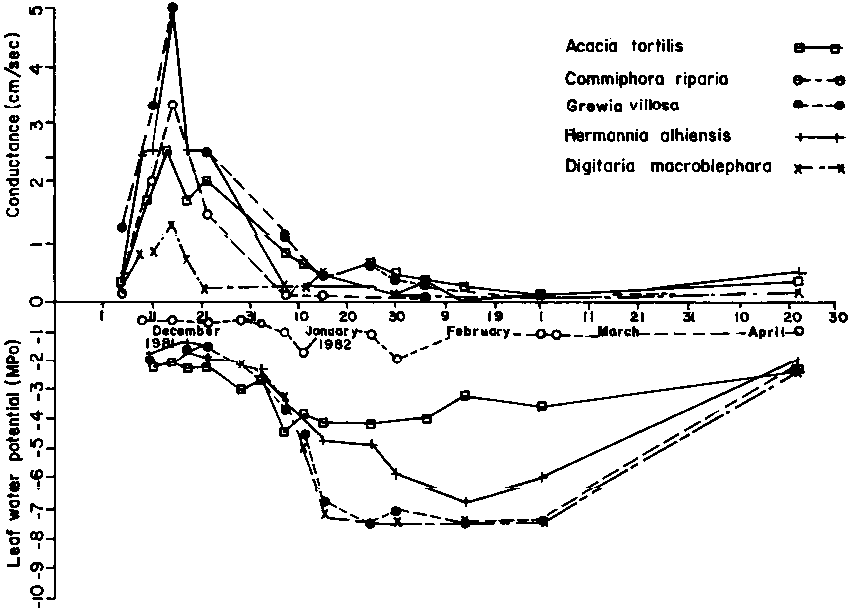
Leaf water potentials for the shrub and herbaceous plant species (see Figure 2) in mid-December were characteristic of non-water deficit conditions. In the course of the wetting phase, leaf water potentials barely dropped below -4 MPa. Thereafter, all other plant species except Commiphora riparia exhibited fluctuating trends of leaf water potentials indicating a cessation of soil water input coupled by soil water depletion to meet transpiration demand. The lowest leaf water potential of -7 MPa was recorded for Digitaria macroblephara and Grewia villosa between January and February. Commiphora riparia maintained leaf water potentials at levels greater than -2 MPa throughout the growth cycle. Leaf water potentials for Acacia tortilis fluctuated between -4 MPa and -1 MPa over the same period. Stomatal conductance exhibited a pattern that was approximately inversely related to leaf water potential at least for the herbaceous plant species studied. Stomatal conductance values peaked in December coinciding with the phase of active soil water replenishment.
Figure 1. Soil water potential by soil depth.
Figure 2. Conductance and leaf water potential by species.
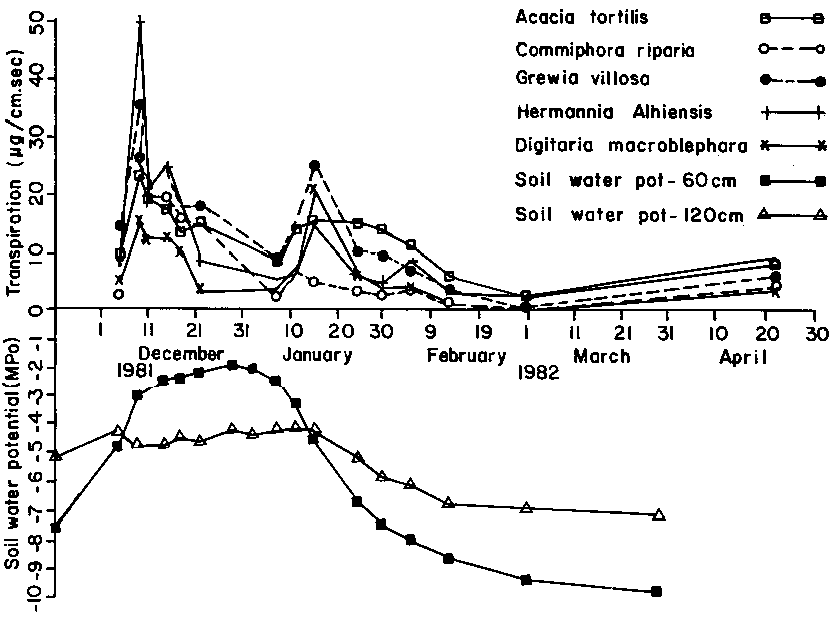
Transpiration rates of the shrub species in the course of the wetting phase ranged between 20 micrograms/cm-2 S-1 and 30 micrograms/cm-2 S-1 whereas that of Digitaria macroblephara rarely exceeded 20 micrograms/cm-2 S-1 (see Figure 3).
Figure 3. Transpiration by species and soil water potential by depth.
In the course of the wetting phase, water was available within the rooting zone and all species maintained a favourable water balance. However, as soil water depletion continued to meet the rising evapotranspiration demand as the dry season advanced, moisture supplies were exhausted from the top soil profiles. Shallow-rooted Digitaria macroblephara and Grewia villosa registered unfavourable water balance. The deep-rooted Acacia tortilis appeared green for the entire growth cycle indicating that water was available within its rooting zone. Commiphora riparia drops most of its leaves and maintains a succulent stem in the course of the dry season. In this study, Commiphora riparia maintained a favourable stable leaf water potential suggesting an efficient regulation of its water economy facilitated by water conservation mechanisms. Feinner (1981) in a study of two Commiphora sp., Adonsonia digitata and Sterculia rhynchocarpa reported that these species are relatively efficient in conserving their water content.
The specificity in water use patterns of semi-arid savanna ecosystems places important restrictions in their responses to management interventions. Excessive removal of herbaceous biomass by grazing animals would influence the total water input reaching the various soil profiles. Walker et al. (1981) reported that on medium to heavy textured savanna soils, decreases in infiltration rates resulting from capping of the soil surface pores were associated with a decline in grass and litter cover.
In most of the Kenyan pastoral grazing lands, excessive depletion of grass cover through overgrazing has resulted in a serious problem of bush encroachment. The environmental processes leading to bush encroachment seem to be closely tied to the differential water use patterns associated with morphological characteristics facilitating exclusive use of water available at different soil profiles.
Soil water potential distribution in the soil profiles of semiarid ecosystems is closely related to the nature of precipitation events and soil water depletion characteristics. The soil moisture response curve is characterised by rapid oscillations in the levels of soil water potential in the top soil profiles. Gradual declines in soil water potential occur with increase in depth and progressive advancement of the dry season.
Shallow-rooted plants like Digitaria macroblephara and Grewia villosa tend to take exclusive advantage of water availability in the top soil profiles when precipitation events occur. As the dry season progresses, these plants quickly show unfavourable water balance suggesting little control of their water economy.
In general the seasonal patterns of transpiration, leaf conductance and leaf water potential suggested that the shrub species utilised more water per unit area than the herbaceous plant species.
I wish to express my appreciation to the technical support staff of the National Range Research Station, Kiboko for assistance in field work. Professional advice rendered by Drs. Dennis Child and M.J. Trlica of Winrock International is sincerely appreciated. This work was supported by funding from USAID and the Government of Kenya.
Feinner, N. 1981. Water loss from cut shoots of arid bushland species in Kenya. Kenya Journal of Science and Technology (B) 2:9-13.
Griffiths, J.F. 1972. Climate. In: W.T.W. Morgani East Africa. Its peoples and resources. Oxford University Press, London
Maranga, E.K., Trlica, M.J. and Smeins, F. 1983. Water relations of Panicum maximum and Digitaria macroblephara on a semiarid rangeland in Kenya. E. Afr. Agric. For. J. 48:74-80.
Maranga, E.K. 1986. An ecological perspective of the phytosociology of woody - Acacia herbaceous - understorey complex. Proceedings of a Conference on Range Development and Research in Kenya, Egerton College, Njoro, Kenya. Winrock International Institute for Agricultural Development.
Michieka, D.O. and van der Pouw, B.J.A. 1977. Soil and vegetation of the Kiboko Range Research station, semi-detailed soil report, No. 53, Draft Edition. Kenya Soil Survey, NAL, Ministry of Agriculture, Nairobi, Kenya.
Pratt, D.J., Greenway, P.J. and Gwynne, N.D. 1966. A classification of East African rangelands. J. Appl. Ecol. 3:369-382.
Sala, O.E., Lauenroth, W.K. and Parton, W.J. 1982. Plant recovery following prolonged drought in a short-grass steppe. Agric. Meteorol. 27:49-58.
Spanner, D.C. 1951. The Peltier effect and its use in the measurement ofsuction pressure. J. Exper. Botany 2:145-168.
Walker, B.H., Ludwig, D., Holling, C.S. and Peterman, R.N. 1981. Stability of semi-arid savanna grazing systems. Journal of Ecology 69:473-498.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}