![]()
![]()
![]()
2.1 Forage intake
2.2 Energy utilization
2.3 Animal reproduction
2.1.1 Ad libitum forage intake
2.1.2 Forage intake in real systems
In general, forage intake by cattle grazing extensively is influenced by the age and physiological status of individual animals, the quality and quantity of the forage on offer, and the environment, including management and climatic factors. Considerable experimental work has been done, particularly in the last two decades, attempting to quantify the separate influences of the individual determinants of intake. This work has suffered from a lack of standardization in the definition of the experimental animals used, the environment of the experiments, the quality and quantity of forage on offer, and above all, a clear definition of the variables being measured. Consequently, it is difficult to synthesize this work into a comprehensible framework; most attempts have only indicated the direction of effects or have achieved quantifications with limited applicability (Belch and Campling, 1962; Baile and Forbes, 1974; Bines, 1976; Cordova et al, 1978).
This section deals first with a synthesis of selected experimental work leading to a specification of ad libitum intake by cattle. An approach is then proposed to modify the specification of ad libitum intake according to the particular environment of the system under study.
Voluntary consumption of forage is limited primarily by the rate of passage of food through the digestive tract. This rate is, in turn, a function of the digestibility of the forage consumed. As a consequence, other factors being equal, digestibility should explain variations in voluntary forage consumption.
Several studies have focused on the extent to which digestibility does in fact explain voluntary forage intake. In grazing situations with a highly variable mix of forage on offer and a high portion of roughage in the diet, the digestibility of the forage is recognized as the predominant factor limiting voluntary food intake (Campling et al, 1962; Blaxter and Wilson, 1962; Conrad et al, 1964; Montgomery and Baumgardt, 1965; Conrad, 1966; Hodgson, 1968; Karue et al, 1973; Baile and Forbes, 1974). As digestibility increases, intake increases substantially, but over a certain level of digestibility (about 65%) voluntary dry matter intake, ceteris paribus, becomes negatively related to digestibility (Conrad et al, 1964; Montgomery and Baumgardt, 1965; Conrad, 1966). Thus, on a diet of highly digestible concentrated feeds, voluntary intake is very closely related to the animal's requirements (Coppock et al, 1974).
Although experiments have shown a strong relationship between voluntary feed intake and digestibility, results cannot be generalized beyond the particular contexts of the experiments. Most of the experiments reported related to mature animals at a certain physiological stage, fed on forage whose digestibility was altered, either artificially through mixing with concentrates or naturally as the growing season progressed.
The usual method of estimating the voluntary forage intake of freely grazing animals is by faecal output (for instance, Conrad et al, 1964; Elliot and Fokkema, 1961; Elliot et al, 1961; Corbett et al, 1963; Hodgson, 1968). Daily intake (I) and daily faecal output (F) are related as
F = (1-d) I ... (2.1)
where d is the digestibility fraction of the forage consumed. Faecal output is usually expressed on the basis of metabolic liveweight, i. e.
F = a W 0.73 ... (2.2)
where a is the rate of passage through the digestive tract (kg/kg metabolic weight/day), referred to as the physical limit, Other things being equal, this rate is a function of the physiological status of the animal (e.g. pregnancy, lactation, etc. Thus, in general
a = f(P) ... (2.3)
where p is physiological status. Equations (2.1), (2.2) and (2.3) solved for I yield:
I = f(p) W 73/(1 - d) ... (2.4)
where:
I = forage intake (kg/day)
W = liveweight (kg)
d = digestibility of forage consumed (fraction)
f(p) = the rate of passage through the digestive tract (kg/kg metabolic weight/day) which is in general a function of the animal's physiological status.
Most reported results fall within this general specification, with differences between the functional forms depending on which variable is held constant. Of studies carried out in tropical Africa, one of the most comprehensive, in terms of number of animal physiological situations analyzed, length of observation period, digestibility range covered and completeness of recorded results, is reported in Elliot and Fokkema (1961) and Elliot et al (1961). Observations of intake were made for a continuous period of 2 years under natural pasture grazing without supplementation. All possible physiological situations were observed, including dry, pregnant and lactating animals. The digestibility of the forage consumed ranged between 42 and 65%, and a highly significant relationship between intake and digestibility was estimated. On the basis of the figures reported by Elliot et al (1961), the derived rate of passage through the digestive tract (coefficient a) for dry cows was 0.042, for cows in the last 3 months of pregnancy it was 0.045, and for lactating cows 0.049. Thus, the intake of pregnant and lactating cows was about 7% and 15% higher than the intake of dry cows. The stage of lactation might also have an effect on intake. Elliot and Fokkema (1961) suggest on the basis of their observations that the lactational stimulus on appetite declines as lactation progresses, so that at weaning the differences between lactating and dry cows are relatively small. However, a comparison by Corbett et al (1963) of intake figures for cows from the 2nd to 4th months of lactation with those for cows from the 5th to 7th months of lactation revealed no apparent relationship between intake and the stage of lactation.
Caution should be used in extrapolating the rates of passage derived from the figures reported by Elliot et al (1961) beyond the digestibility limits of 42 to 65% within which they experimented. Very low quality forage, for instance with a crude protein content below 6% which corresponds to approximately 40% digestibility (Grover et al, 1957), substantially depresses forage intake. To account for this effect, Sanders and Cartwright (1979), drawing from results reported by Campling et al (1962), assumed an additional multiplicative adjustment in voluntary intake for crude protein (CP) less than 0.06 (6%) by the factor (CP/0.06)0.6. Assuming a close relationship between crude protein and digestibility, this adjustment can be expressed in terms of digestibility alone, defined as (d/0.4)0.6, 6, being in effect when digestibility is below 40%.
Similarly, as digestibility rises above about 65%, chemostatic or thermostatic mechanisms appear to regulate intake (Montgomery and Baumgardt, 1965; Conrad et al, 1964; Conrad, 1966; Baile and Forbes, 1974), implying, ceteris paribus, a constant energy intake for these high digestibility levels. Thus, it will be assumed that when the digestibility of forage is greater than 65% the animal's feed intake will be adjusted to maintain metabolizable energy equal to the level at 65% digestibility, as implied by equation (2.4):
d. I = 0.65 f(p) W 0.73/(1 - 0.65)
which yields
I = 1.86 f(p) W 0.73/d ... (2.5)
This constraint to voluntary intake at high digestibility levels is referred to as the physiological limit.
In addition to the quality of the forage on offer and the size and physiological status of grazing animals, the level of ad libitum intake is also a function of the animal's age. Very young calves (1 to 2 months) consume insignificant quantities of forage as they cannot digest a greater intake. The digestive system of ruminants develops gradually, reaching adult capacity at about 18 months.
Relatively few quantitative estimates of voluntary forage intake by young animals are reported in the literature. Hodgson (1968) experimented with calves from 3 to 6 months, grazing on forage of digestibility ranging from about 65 to 80%. Intake was linearly related to digestibility within this range, indicating that the chemostatic mechanisms which limit intake in mature animals are not operating in young, rapidly growing calves. The relationship obtained was:
I = (- 0.143 + 0.3d) W 0.73
where:
I = daily voluntary intake (kg dry matter)
d = digestibility (fraction)
W = liveweight (kg).
Information is apparently lacking on forage intake by young animals at low digestibility levels. An extrapolation of the above function implies that intake would be zero for digestibility below about 48%, which indicates that extrapolation downwards for the full range of digestibilities encountered is not possible. However, Hodgson also reports information on faecal output which provides an indirect measure of intake. He found that daily faecal output had an upper limit of about 22 grams per kg metabolic weight. Thus, voluntary intake by calves can be expressed by the equation (2.4), where coefficient a has the value 0.022 or about 53% of the coefficient obtained for dry cows.
Similarly, although it is recognized that the voluntary intake of old animals is reduced due to loss of teeth and other problems associated with aging, quantified data are lacking. Sanders and Cartwright (1979) assume that voluntary intake is reduced by 3% for each year of age beyond 8. Thus, for animals older than 8 years, voluntary intake as reported earlier is adjusted in a multiplicative fashion by the factor (1 - 0.03 (t - 8)), where t is age in years.
In summary, ad libitum intake by freely grazing cattle can be expressed as a function of the quality of the forage on offer and the age, size and physiological status of individual animals. However, it would be incorrect to apply the absolute levels of the intake coefficients as reported above to all production systems. Other factors, such as the palatability of the forage, ambient temperature, regularity and quantity of water on offer and breed adaptability, have not been explicitly included. However, in relative terms, the intake coefficients for various classes of animals should be comparable between different production systems. Thus, as explained later in Section 3.3.1, the intake coefficient for a reference class of animals is estimated, based on observations from the system under study, and from this estimation the intake coefficients for other classes of animals are obtained.
The estimates of forage intake reported above assume an ad libitum feeding regime and also adequate time for grazing. In many systems intake might be reduced because the quantity of forage on offer is limited, or daily grazing time might be restricted due to environmental constraints (such as heat) or considerable time spent walking.
Greenhalgh et al (1966) studied the effect of grazing intensity on herbage consumption and estimated that about 60% excess feed should be available for maximum consumption to be achieved. Similarly, Zemmelink (1980) studied the effect of selectivity on voluntary intake and estimated that 15 to over 40% excess feed was required to achieve maximum intake, depending on the quality of the forage. The general model suggested is:
where:
y = level of voluntary intake
x = quantity of forage on offer
m = upper limit for y
p = fraction of the forage which may be considered edible or acceptable
h = a parameter indicating the shape of the relationship between x and y.
This specification assumes no limitation on the time available for feeding and thus might not apply in many situations. Wilson and Flynn (1974), for example, report that beef cattle offered grass silage ad libitum required at least 6 hours access for maximum intake. Access times for extensively grazing cattle would be expected to be much higher for maximum intake, depending on the spatial concentration of the forage on offer. Thus, a more complete specification determining actual intake should take into consideration both quality and quantity of forage on offer and the time during which animals have access to this forage. The following is an attempt to provide such a general specification, starting with the assumption that the rate of forage consumption per unit of time is a function of the quantity of the forage on offer, provided the forage quality is adequate and the forage can be consumed by cattle:
r = f (Q1) ... (2.6)
where:
r = rate of forage consumption (kg dry matter/hour)
Q1 = quantity of consumable forage on offer acceptable to the grazing animals (kg dry matter/ha).
It should be expected that
That is, the rate of consumption is higher, the higher the quantity of acceptable forage on offer. The general shape of the relationship between r and Q1 is depicted in Figure 2.1.
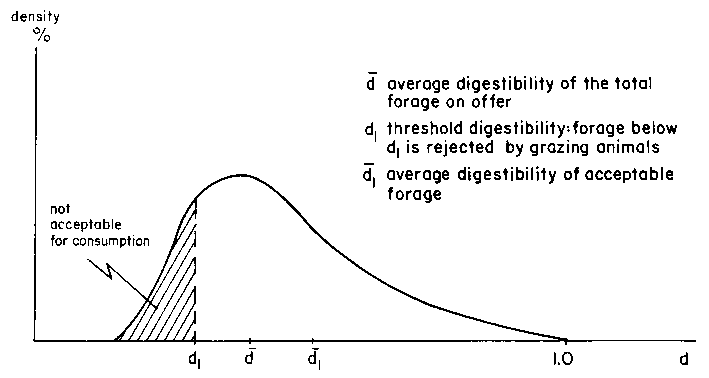
Whether the level of voluntary intake is achieved as estimated from the digestibility fraction depends on the quantity of acceptable forage on offer and the length of grazing time allowed in the system under consideration. The forage on offer will be characterized by a quality distribution function as shown in Figure 2.2 which provides the relative quantities of forage of different digestibilities. From this distribution function, it is possible to obtain a forage supply curve as shown in Figure 2.3 relating quantities of forage on offer above a given digestibility level. Thus, if d is the average digestibility of the total forage on offer (QT) and if the total forage is consumed, then its digestibility will be equal to d.
Figure 2.1 Rate of consumption (r) in kg/hour as a function of the density of acceptable forage on offer (Q1) in kg/ha
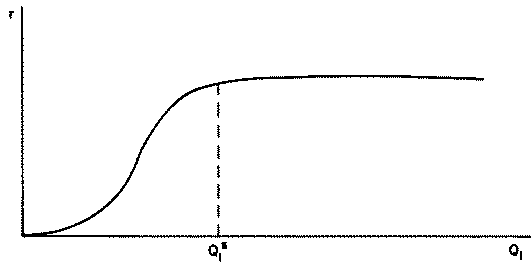
Figure 2.3 Forage supply curve: relationship between quantity of forage grazed (Q) and its average digestibility (d)
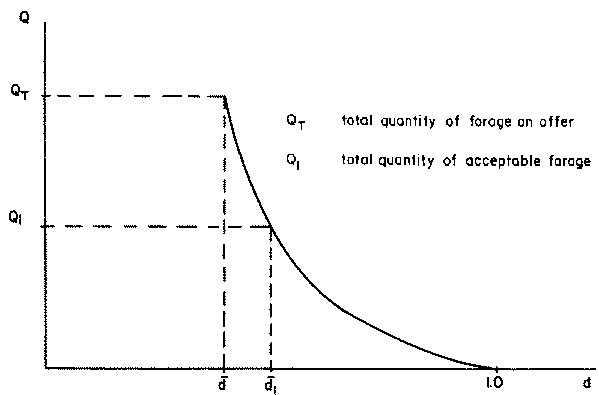
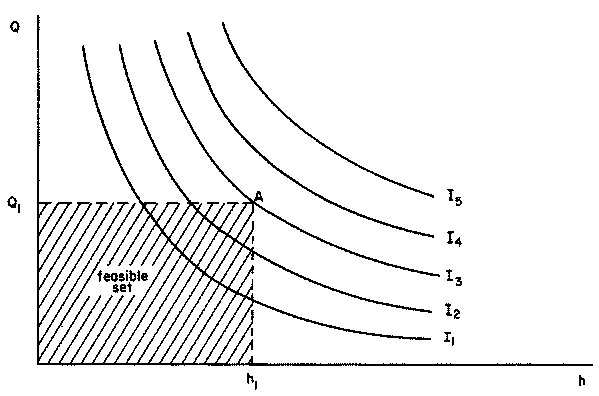
Figure 2.4 Isointake curves: relationship between quantity of acceptable forage on offer (Q) and length of grazing time (h) for different levels of intake (I1, I2, ...)
In general, animals will reject forage below a certain minimum digestibility (d1) which implies an average digestibility of acceptable forage 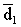 , and a corresponding quantity Q1, such as
, and a corresponding quantity Q1, such as  and Q < QT.
and Q < QT.
Given relationship (2.6) between the rate of consumption and the density of the forage on offer, the total intake during h hours of grazing time will be
I = r. h = f(Q1).h ... (2.7)
where:
I = total intake (kg)
h = grazing time (hours).
This relationship suggests a trade-off curve between the quantity of forage on offer in a given area and the length of grazing time. A given intake level can be obtained from dense forage in a short time or from less dense forage over a longer period. These trade-off curves will be called isointake curves, representing the different combinations of forage on offer and length of grazing time that will yield the same intake. Figure 2.4 shows a family of isointake curves for different intake levels. In any real situation, the time actually available for grazing (h1) is limited by management and environmental factors, while forage on offer is limited to the quantity of a minimum acceptable digestibility level (Q1). Thus the feasible set of forage quantity and grazing time is shown in the shaded area of the figure.
Thus, the level of voluntary intake as estimated from the digestibility fraction is achieved if its corresponding ad libitum isointake curve crosses the feasible set of Figure 2.4. The maximum actual intake corresponds to the level of the isointake passing through point A. Conversely, if the ad libitum isointake does not cross the shaded area, then actual intake is less than the estimated voluntary intake.
2.2.1 Maintenance
2.2.2 Lactation
2.2.3 Pregnancy
2.2.4 Liveweight gain
2.2.5 Energy released from mobilization of body reserves
2.2.6 Energy available to calves from milk consumed
Organic nutrients obtained from the different sources of feed available to an animal are used for a variety of purposes, including the maintenance of body functions, the construction of body tissues, the synthesis of milk, and the conversion to mechanical energy used for walking and other work. These diverse functions all require the transfer of considerable quantities of energy, so that in most situations when the energy requirements of the animals' different functions are met it may be assumed that the animal's non-energy requirements (protein, minerals and vitamins) are also met. Hence, the nutritive value of different feeds can be expressed by their energy content, or more precisely by their ability to supply energy with a high coefficient of conversion into usable energy for the different body functions.
The gross energy contents of different forages are very similar, at about 18 MJ/kg (Hunt, 1966). As shown in Figure 2.5, a portion of this energy is lost as faeces, while the remaining digestible energy (DE), proportional to the digestibility (d) of the consumed feed, is converted into metabolizable energy (ME) after additional losses of about 19% of DE as urine and methane (Armstrong, 1964; MAFF, 1975). Metabolizable energy expressed as a fraction of gross energy is referred to as metabolizability (q), and relates to the digestibility of the consumed forage approximately as:
q = 0.81 d ... (2.8)
Thus, metabolizable energy obtained from a given intake can be expressed as:
ME = 18I. q ... (2.9)
or equivalently
ME = 14.6 I. d
where:
ME = metabolizable energy (MJ)
I = feed intake (kg)
d = digestibility of feed intake (fraction).
The conversion of metabolizable energy into net energy is also associated with some losses as heat, which depend on the quality of the consumed feed and the body function for which the energy is utilized. In the most general case, energy use in an animal can be accounted for by its requirements for maintenance, lactation, pregnancy and growth. Thus, conversion of metabolizable energy to net energy is described by the relationship:
Ei = ki . ME = 14.6 ki . Ii . d ... (2.10)
where:
Ei = net energy (MJ) required for the i'th body functionki = coefficient of the efficiency of conversion of metabolizable into net energy for the i'th body function
Ii = quantity of feed (kg) required to meet the energy requirements of the i'th body function
d = digestibility of consumed feed.
Solving (2.10) for Ii provides the quantity of forage needed to meet the energy needs of the i'th body function:
Ii = Ei/(14.6 ki . d) ... (2. 11)
Thus, for the calculation of intake required for the i'th body function, net energy requirements (Ei) and the efficiency coefficients (ki) need to be specified. This is done in the next sections, followed then by formulae specifying the net energy released from the mobilization of body tissue in the case of inadequate feed intake and the energy available to calves from milk consumption.
Maintenance can be defined as that state of the animal in which there is neither a net gain nor loss of nutrients (Kay, 1976). Maintenance requirements are estimates of the amount of nutrients required to achieve such an equilibrium. One component of the energy requirements for maintenance is referred to as basal metabolism, and is proportional to the metabolic body size of the animal. The second component of the energy requirements for maintenance relate to the level of the animal's activity and can he expressed approximately by liveweight and the daily distance walked. Thus, following Blaxter (1969) and Webster (1978), total net energy requirements for maintenance can be obtained from the relationship:
Em = 0.376 W 0.73 + 0.0021 W.D ... (2.12)
where:
Em = net energy requirements for maintenance (MJ/day)
W = liveweight (kg)
D = distance walked (km/day).
Figure 2.5 The partition of feed energy in an animal (adapted from McDonald et al, 1978)

The efficiency with which metabolizable energy is used for maintenance (km) can be expressed as a function of the metabolizability of the consumed forage (see, for example, Blaxter, 1974; van Es, 1976; MAFF, 1975; Pigden et al, 1979):
km = 0.55 + 0.3 q ... (2.13)
Substituting. the values of (2.12) and (2.13) into equation (2.11) yields the feed requirements needed to meet energy for maintenance. In general, the quantity of feed required increases for heavier animals and greater activity and decreases for feed of higher quality.
Net energy requirements for lactation are approximately proportional to the quantity of milk produced:
E1 = e1 M ... (2.14)
where:
E1 = net energy requirements for lactation (MJ/day)
e1 = energy content of milk (MJ/kg)
M = milk yield (kg/day).
The energy content of milk is approximately given by the relationship (MAFF, 1975):
e1 = 0.0386 BF + 0.0205 SNF - 0.236 ... (2.15)
where:
BF = butter fat content (g/kg)
SNF = solids-not-fat content (g/kg).
For example, for a BF of 54 g/kg and an SNF of 85 g/kg, which approximately corresponds to milk from Zebu cattle (Williamson and Payne, 1978), the calculated energy content is 3.6 MJ/kg.
The efficiency with which metabolizable energy is converted into net energy for lactation (k1) is less than that for maintenance. The suggested relationship (Pigden et al, 1979) is:
k1 = 0.463 + 0.24 q ... (2.16)
The significance of the metabolizability coefficient is not very high (van Es, 1976), and a value for k1 of about 0.60 is usually suggested (e.g. MAFF, 1975).
Substituting the values of (2.14), (2.15) and (2.16) into equation (2.11) yields the feed requirements to produce a given quantity of milk of a certain energy content. In general, the quantity of feed required increases for higher milk yields and higher energy contents and decreases for feed of higher quality.
Net energy requirements for pregnancy involve the energy deposited in the uterus and associated tissues (Ep1), the energy associated with synthetic processes involved in foetal growth (Ep2), the energy required for foetal maintenance and the increased maternal fasting metabolism occurring during pregnancy (Ep3). Thus total net energy for pregnancy as given by MAFF (1975) is defined as:
Ep = Ep1 + Ep2 + Ep3 ... (2.17)
where:
Ep1 = 0.03 e 0.0174t
Ep2 = Ep3 = 0.452 e 0.1t
t = number of days after conception
e = base of the natural logarithms (approximately 2.718).
The efficiency with which metabolizable energy is converted into net energy for pregnancy depends on the different end uses of this energy. Thus, associated with the three net energy expenditures as described above there are corresponding efficiency coefficients as follows (MAFF, 1975):
kp1 = kp2 = 1.0 ... (2.18)kp3 = km
where:
km = the coefficient of the efficiency of converting metabolizable energy m into net energy for maintenance given by equation (2.13).
Net energy requirements for liveweight gain depend in general on both the liveweight gain achieved and the animal's present liveweight. A relationship suggested by MAFF (1975) is:
where:
Eg = net energy requirements for liveweight gain (MJ/day)
DW = liveweight gain (kg/day)
W = present animal liveweight (kg),
Conversely, the liveweight gain obtained from the availability of net energy Eg is given by the inverse of the above relationship, i. e.
DW = Eg/(6.28 + 0.0188W + 0.3 Eg) ... (2.19)
The efficiency with which metabolizable energy is converted into energy for weight gain (kg) is lower than that for lactation and is given approximately by the relationship (Blaxter, 1973):
kg = 0.03 + 0.81 q ... (2.20)
However, for liveweight gain during lactation, kg is much higher and approximately equals the efficiency coefficient for lactation:
kg = k1 ... (2.21)
or is even higher, according to experiments by Moe et al (1971).
Substitution of relationships (2.19) and either (2.20) or (2.21) according to pregnancy status into relationship (2.11) yields the feed requirements for live weight gain. In general, the quantity of feed required increases for higher liveweight gains and heavier animals and decreases for feed of higher quality.
When energy derived from feed intake is insufficient to meet the animal's total energy requirements, then the energy balance is achieved by the mobilization of body reserves. This is common soon after parturition, but in situations of low quality and quantity forage on offer and without adequate supplementation an animal may need to mobilize body reserves to meet pregnancy and maintenance requirements.
Body tissue has an energy value of about 20 MJ/kg. Energy released by tissue mobilization is lower than the energy that originally went into its synthesis. Moe et al (1971) obtained the following relationship for energy utilized for milk production and originating in body tissue loss:
E1 = 0.632 ME - 0.84 TL - 79.7
where:
E1 = net energy deposited as milk (Kcal/kg metabolic weight)
ME = metabolizable energy (Kcal/kg metabolic weight)
TL = tissue loss energy (Kcal/kg metabolic weight).
This expression indicates that the coefficient of conversion of energy from body tissue mobilization to net energy for lactation is 0.84. MAFF (1975) suggests a value of 0.82. Thus, for example, if 10 kg of milk are produced, with an energy content of 3.6 MJ/kg, drawing exclusively on body tissue mobilization, this would imply a weight loss of 2.19 kg computed as (10 kg x 3.6 MJ/kg)/(0.82 x 20 MJ/kg). The same efficiency coefficient is assumed when energy released from body tissue mobilization is used to meet pregnancy or maintenance requirements.
In general, assuming an energy value of 20 MJ/kg for body tissue and a coefficient for its utilization of 0.82, the net energy available from the mobilization of body tissue is:
E = 16.4 DW ... (2.22)
where:
E = net energy available for maintenance, pregnancy or lactation (MJ)
DW= mobilized body tissue (kg).
The amount of body tissue needed to meet a net energy deficit E is given by the inverse:
DW = E/16.4 ... (2.23)
Milk consumption is the main source of energy for young calves. The gross energy content of milk is given by equation (2.15) as a function of butter fat and solids-not-fat content. The fat fraction of milk accounts for about 60% of its energy content and is highly digestible (almost 98% for young lambs) (Owen, 1976). The overall digestibility of cow milk is nearly 93% (MAFF, 1975), which implies a digestibility level of around 86% for its solids-not-fat content. Metabolizable energy from milk is also used with great efficiency, at about 75% for lambs (Owen, 1976).
Based on these conversion factors, and assuming a close correspondence between lambs and calves, the net energy derived by calves from milk consumption is about 70% of the gross energy contained in the consumed milk (0.93 × 0.75 = 0.70). Thus the net energy available from a given quantity of consumed milk is given by:
E = 0.70 e1 M ... (2.24)
where:
e1 = gross energy of milk (MJ/kg) as given by equation (2.15)
M = milk consumption (kg).
The quantity of milk needed to meet a net energy requirement is given by the inverse:
M = 1.43 E/e1 ... (2.25)
Reproduction in a cattle herd is a complex phenomenon. A number of factors have been identified in the literature as determining the rate of reproduction of individual females in a herd, including age, liveweight level and change, and post-partum period (Lamond, 1970; Buck et al, 1976; Newton et al, 1980; Pleasants and Ginindza, 1980).
In estimating reproduction rates, the females in a herd are classified in two categories, heifers and mature cows. The timing of a heifer's first conception is a major factor influencing its lifetime productivity, and is usually subject to some control by management to ensure that conception does not occur when an animal is in poor condition and that subsequent calving will occur at an appropriate time. Primarily, however, the timing of the first conception is determined by age and liveweight (Lamond, 1970; Pinney et al, 1972; Siebert and Field, 1975; Drennan, 1977; Anderson, 1977). According to these and other studies, puberty is reached at an average liveweight which is largely genetically determined, though the standard deviations for age and liveweight at puberty are large. If animals are inadequately nourished, their growth rate is reduced and the onset of puberty is delayed (Joubert, 1963). This is particularly evident in harsh environments where nutrition levels for young stock may be seriously inadequate after weaning. In such cases, heifers may need to reach higher weights than normal for their breed at the time of first conception (Sparke and Lamond, 1968). The variability in age or liveweight at puberty is probably greater than normal in undernourished animals (Lamond, 1970). At the other extreme, heifers which have grown very rapidly tend to conceive less readily than heifers which have grown at a moderate rate (Reid et al, 1963).
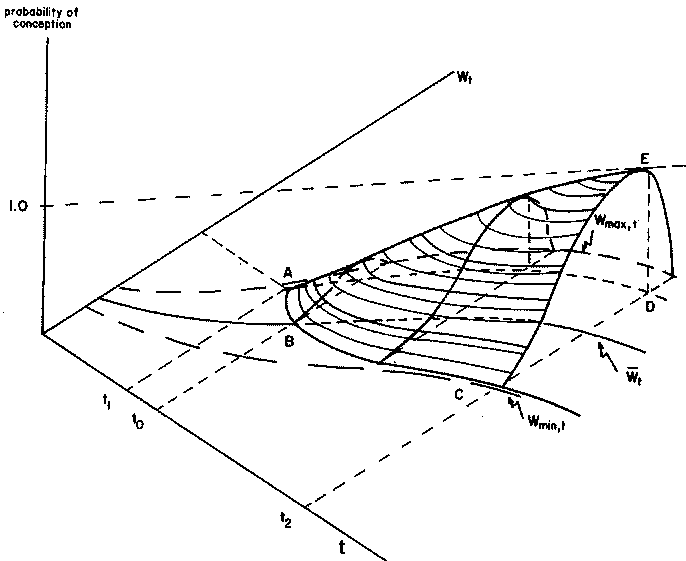
Assuming that age and liveweight account for most of the variability in heifer conception rates, then the above observations lead to the construction of a probability surface representing heifer conception, as presented in Figure 2.6. The relationship represented in this figure has the following features:
1. There is a minimum age/liveweight combination at which conception can occur (point A).2. There is an 'ideal' liveweight at any age with a maximum probability of conception (segment (AD', the projection of the segment (AK) onto the liveweight-age plane).
3. The non-zero probabilities of conception correspond to weights above a minimum liveweight boundary (Wmin, t) and below a maximum liveweight boundary (Wmax, t) characteristic of the breed and the production system.
4. The surface is regular and smooth. Thus, it permits the derivation of isoprobability of conception curves, defining liveweight/age combinations with equal probabilities of conception.
Such a formulation of the determinants of conception in heifers is well suited to use in a computer simulation model. The use of this formulation in regard to the other components of the model is discussed in detail in Section 3.3.3.1.
For mature cows after first calving, conception rates are basically a function of age, with maximum calving rates achieved in the middle age range and lower observed conception rates for both younger and older animals. The general age effect on average conception rates is shown in Figure 2.7. Such relationships are usually derived from an aggregation of data from several years (e.g. Buck et al, 1976). While they usually explain a high portion of observed fertility for different age groups, some unexplained variance arises principally from the particular management system used and genetic differences between animals. Shifts upwards or downwards in fertility levels would be expected for herds with higher or lower average planes of nutrition. Buck et al (1976) show such a curve for the total animal population observed, but not for each age class. Their results show substantially higher calving rates for cows in good condition relative to those in poorer condition. At the extreme, however, the reproductive performance of very heavy cows is marginally lower than that of cows in average to good condition.
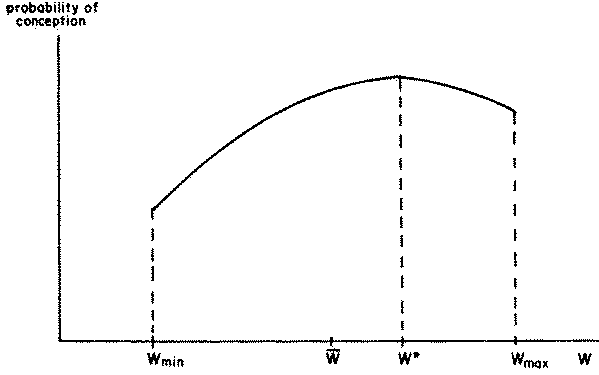
When sufficient data on conception rates exist and are disaggregated by age and liveweight, a family of curves can be obtained, with each curve corresponding to a given age class. An exemplary curve, for a given age class, is shown in Figure 2.8. The relationship represented in this figure has the following features which are common to all age classes:
1. The curve is truncated on the left at liveweight Wmin and on the right at liveweight Wmax representing the lowest and highest liveweights observable for this age class.2. Between these limits, the curve is smooth and continuous.
3. The maximum conception rate corresponds to an 'ideal' liveweight for conception, intermediate between Wmin and Wmax and, in general, closer to the upper limit of liveweight.
Figure 2.6 Liveweight/age effect on the probability of conception of heifers
Figure 2.7 Effect of age on cow fertility
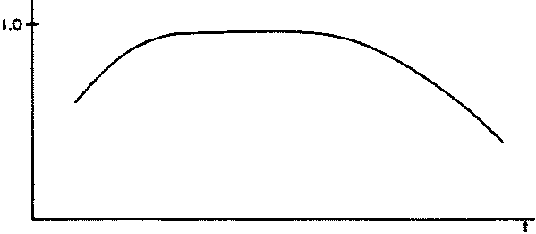
Figure 2.8 Effect of liveweight on conception
Conception in cows is also influenced by the length of the post-partum period. Conception virtually never occurs in the first month post-partum, but subsequently this effect diminishes rapidly. It is not usually considered significant by the 2nd or 3rd month post-partum.
![]()
![]()
![]()
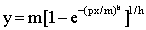
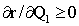
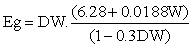
{kind=link}
{kind=link}