![]()
![]()
![]()
Modern strategies of livestock production require exogenous inputs of skills, equipment and money. A fundamental element of these strategies is the new ability to place water points where one wants them to be, rather than where they occur by accidents of nature. Unfortunately these technological innovations are entirely dependent for their success on the human organisation and management of livestock and grazing resources. In pastoral systems, where the traditional organisation has collapsed and has not been replaced by an alternative, overgrazing becomes a certainty and the provision of extra water a liability. However, water development makes such an immediate improvement to the quality of life of pastoral people that its provision can hardly be denied. Having accepted the social and political obligations to provide water, the emphasis must be on minimising its deleterious effects.
In theory, the objective of water development in pastoral areas is similar to that for commercial ranching areas, but in practice it is quite different because of this lack of grazing control and the resultant range degradation around watering points. The extent of this degradation has been described by Jarrige (1980) relying largely on information from Le Houérou (1977):
"The quality, productivity and surface of the arid and semi-arid pastures of Africa (and the Middle East) have deteriorated at an alarming rate over the last decades as a result of overstocking and the increasing pressure of human population. Production is currently one third to one fifth of its potential and sometimes even as low as one tenth in the arid zone of Africa. The numbers of livestock have increased sharply (38% in all of Africa between 1950 and 1973; 53% for cattle) as a result of better sanitary protection and increased rainfall in the 1950s and 1960s, sometimes helped by unsound water development projects. Heavy and continuous overgrazing have resulted in the replacement of perennial grasses by annual grasses and of forage shrubs and trees by undesirable ones. At the end of the 1968-1973 drought in the Sahel, the vegetation cover was 32% less than 30 years ago. Tens of thousands of hectares are converted to desert by wind erosion every year. The human population explosion has led to more and more pastures being turned into cereal cropping and fallow being reduced or suppressed. This periodical cereal cropping with modern ploughs eliminates the perennial forage species. Destruction of woody species for fuel around the cities not only suppresses some browse species but also results in the disappearance of valuable forage species that grow in the shade."
The prospects for the future are not encouraging. Food problems in tropical countries are well known. The rise in food production will have to exceed population growth in order to remedy present-day shortages, which cause half a billion people to suffer from undernutrition or malnutrition. There is little chance of this happening in the near future, because the population has been growing and continues to rise at an unprecedented rate (>2% p.a.). This population explosion is expected to last several more decades. In the intertropical zone by the year 2000 the population will probably reach 2950 million people, or 47% of the world's population and 60% more than at present (Jarrige, 1980).
If pastoralism in any form is to survive, the range resources must be protected above all else.
The most conspicuous effects of range degradation are to be found around permanent wells and boreholes. For example the man-made desert round the settlement at Kom in northern Kenya is nearly 8 km in radius and 200 km2 in area (IPAL, 1982). In Niger, Rippstein and Peyre de Fabregues (1972) estimated that proper exploitation of a circle of 8 hen radius around a pumping station, with overexploitation during 9 months of the dry season, allowed a maximum of 3000 TLU on pastures having an annual primary productivity of 700 kg DM.ha-1. The actual biomass of stock was usually more than 10 000 TLU. The authors made a number of standard recommendations on how water could be used to improve range management. The system of transhumance was to be maintained and the time spent in the wet season dispersal area was to be increased by increasing the number and depth of temporary ponds. The early return of transhumant herds to dry-season grazing grounds was to be discouraged by delaying the date at which the borehole pump starts operating. The number of pumping hours per day were to be varied to deliver the daily water needs for the correct stocking rate. This daily ration was carefully calculated for each month, and varied from 24 l in January to >42 l in October. In addition, dry-season grazing reserves were to be created particularly from wooded pastures and firebreaks were to be introduced. Control of permanent water points, stocking rates and livestock distribution was to be placed in the hands of the nomads who traditionally use the area.
However, it should be noted that the effectiveness of such plans can be limited by: isolated and sporadic rainstorms which may not fill all the ponds in the wet season; pump attendants and range management officials may be bribed or coerced into operating pumping stations (Sandford, in press); the committees set up to manage the rangeland may not be very effective; and fuel delivery and maintenance of pumps and engines in remote areas may be difficult at best.
Further water development in areas of uncontrolled grazing may temporarily alleviate overstocking. However, it will merely postpone the day of reckoning when the rangeland will be a desert unless limited extent of range management in Africa is recognized. The emphasis in water development must be on the continuing use of traditional watering practices for which the labour and social organisation required act as a constraint on range utilisation (Helland, 1980). An example of such a development plan is given by King et al (1983) using carefully spaced hand-dug wells. The preference for wells rather than ponds was based on their reliability and water quality, compared with the unreliability, high evaporation rate, as well as erosion, silting and polluted water, associated with ponds (IPAL, 1982).
In certain situations grazing is controlled. For example, in parts of Kenya Maasailand with an annual rainfall >600 mm individual ranches have been created which are a natural extension of the Maasai practice of reserved grazing areas. Most households have reserved grazing areas for their calves (called olepolole), and some have reserved areas for grazing in the early to mid dry season (dokoya unkishu), as well as reserves (enkaroni) for the final period of the dry season. Households associated with a specific reserved area do not have exclusive rights to that area, but they do collectively decide when livestock may enter for grazing. A meeting of men from a particular neighbourhood makes this decision, and disputes about entry often occur. Nevertheless, collective action may be taken against any person herding prematurely in the reserved area, whether they are from the controlling neighbourhood or from elsewhere (Peacock et al, 1982).
In such cases, water development should be largely a question of economics, which means that a watering point would be required to support the equivalent of 1250 adult zebu cattle (Classen et al, in press). Conventional range management practices may be applied (Pratt and Gwynne, 1977), but these are unlikely to include expensive improvement of natural pastures until research has shown that new interventions (e.g. forage legumes) are a realistic proposition. Food and mineral supplements can be considered, provided they are cheap enough. For example, the provision of urea when forage digestibility is low has increased the milk yield of Merino ewes (Stephenson et al, 1981).
Consequently the protein and fibre content of the diet may place a ceiling on ruminant DMI particularly by bulk grazers such as cattle. Crude protein levels of less than 7% markedly reduce the intake of natural grasslands in the tropics (Osbourn, 1976), and are considered the major limiting factor to liveweight gain, for example in Botswana (Pratchett et al, 1977). Therefore one can apply the old adage that 'water can be limited when forage is short' (in quality or quantity).
Earlier in the dry season, water should be accessible enough to allow the lactating cow to satisfy its appetite and maintain energy balance. That situation is illustrated in Table 46 where the lactating zebu cow is in energy balance, walking about 9 km.d-1. The distance itself is not critical because the water and energy costs of walking are relatively small. Long distances (estimated at >14 km by Konandreas and Anderson, 1982) are nevertheless associated with a loss of grazing time, which is probably a more important factor. Daily watering is necessary for maximum productivity when grass digestibility is above 50% but moisture content below 50% or thereabouts. The conclusion reached is that doubling the density of watering points in an underdeveloped area produces a spectacular improvement in productivity, but that there is a rapid diminution in the return with each doubling in density thereafter (Sandford, in press).
The contribution of trees, particularly acacias, to pastoral systems is considerable. Besides providing green leaves for browsing, all the litter which falls on the ground, in the form of dead leaves, flowers and seedpods, is eaten. Dead wood is used for firewood and live branches for bomas. This last practice results in the destruction of woodland in the vicinity of bomas. Tree regeneration is prevented by overbrowsing particularly by goats which should be excluded from an area for 5 years to allow young trees to grow out of the animals' reach (IPAL, 1982). The destruction is exacerbated by the likelihood of traditional watering places being located where ground water is at or near the surface and trees are concentrated. Careful spacing of new watering points away from some areas of woodland may reduce destruction, but is unlikely to eliminate it because smallstock can forage up to 20 km and bomas can be located up to 15 km from water.
Tree shade provides a shelter for nutritious plants and the resultant green vegetation acts as a heat sink for radiant heat from the animal. Its value for pastoral production in semi-arid rangelands is unquestioned but unquantified. It may be less important for indigenous stock, such as Boran cattle, in more humid less water-limited environments but tree shade is needed to maintain high levels of production if there are high solar heat loads (Fuquay, 1981). Thus shade from mature coconut palms significantly increased the milk yield of Friesian × Boran cows (+18%) on the Tanzanian coast (Macfarlane and Stevens, 1972).
6.3.1 Beef versus milk
6.3.2 Cattle improvement
6.3.3 Breeding cattle for pastoralism
6.3.4 Species mix
6.3.5 Centripetal watering
6.3.6 Restricted watering
6.3.7 Night grazing
Water development projects which take into account the need to minimise range degradation from uncontrolled grazing will clearly need to limit the number and size of water points. The attributes of indigenous livestock in energy and water conservation show them to be better suited to such pastoral production systems than are exotic breeds and crosses.
The main reason for the existence of 100 million of the 135 million cattle in sub-Saharan Africa (FAO, 1977) is to provide a staple diet of milk under conditions which are harsh for both man and beast for much of the year. The amount of milk produced is, of necessity, only a few litres per day but is usually shared between the household and the calf, so that there is scope for marketing immature animals. Furthermore, the high percentage of females in the herd (55 to 60%) provides scope for rapid population recovery after a drought (King et al, in press).
The dependence of pastoralists on milk may be decreasing with the trend towards smallstock, dryland farming and a more cash-oriented economy. Nevertheless, the transition from pastoral milk to commercial beef production is not the logical extension of this trend because it fails to provide a livelihood for the vast majority of the pastoral community. The slums and shanty towns around most African cities bear witness to the hopelessness of trying to find alternative employment for these people.
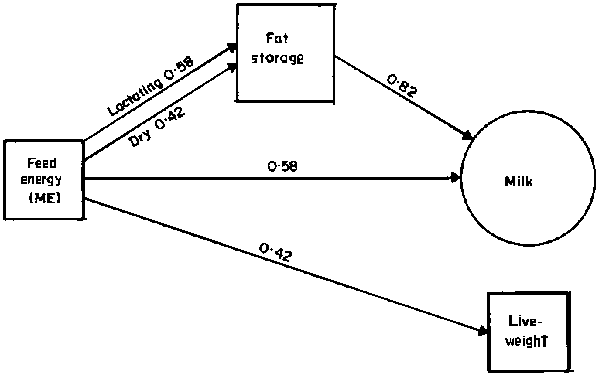
Furthermore, there is nothing inefficient about dairy ranching versus beef. The value, in terms of human nutrients, of a kg of milk is much the same as that of 1 kg of liveweight gain. The efficiency of conversion of feed energy (ME) to milk is much higher than to meat and fat. Lactation also enhances the conversion efficiency of fat deposition. The actual conversion coefficients vary with the digestibility of the diet, which has been fixed at 60% for the purposes of the example (Figure 23). The comparative water content may also be relevant in the dry season when water intake is limited. At that time fatty tissue, containing only 0.451.kg-1, is unlikely to be deposited and the water content of milk (0.87 l.kg-1) and lean meat (0.75 l.kg-1) are quite similar.
The African cattle breeds which are most frequently improved are zebu and Sanga. Compared with European breeds, zebu cattle are generally more tolerant of the heat, parasites and diseases of the African environment. They require less water per kg bodyweight and per degree rise in ambient temperature, but this is mainly due to a smaller food intake and a lower metabolic rate. The threshold ambient temperature at which they start to sweat is higher (28°C as opposed to 17°C), but they sweat at a higher rate once they have started (Winchester and Morris, 1956; Horrocks and Phillips, 1961; Brown and Hutchinson, 1973). Zebu cattle digest low-quality grass hay slightly better than Herefords and show less depression of appetite when water is restricted (Phillips, 1960; 1961a; 1961b). The N'Dama, an indigenous Bos taurus breed found in the humid areas of West Africa, is reputed to have an even lower water requirement than the zebu (Pagot, 1974).
Figure 23. Relative efficiencies of metabolisable energy for cattle production, at a digestibility of 0.60. (Source: Moe and Flatt (1969); Konandreas and Anderson (1982); Van Soest (1982)).
The improved breeds are usually exotic to Africa and come from Europe, North America or Asia. Their introduction is often not questioned because, at the level at which decisions are made, both the donor and the recipient are familiar with the exotic animal in its original environment. For example, attempts to establish the Red Sindhi × Jersey as a dairy breed in the southern United States were abandoned in favour of existing European breeds (Branton et al, 1966). The reason was that it was more economic to ameliorate the environment with cool buildings and reduce the heat increment of feeding with a low fibre, high protein and energy ration than replace high yielding European cattle with more robust zebu crossbred cattle of lower productivity. Since the early days of domestication, 'invaders have persisted with the introduction of livestock to which they are accustomed, even when the habitat was inappropriate' (Bökönyi, 1969).
In many parts of the tropics, for example in Queensland and East and central Africa, there is only limited opportunity for improving the environment and zebu crosses have proved superior to pure European breeds (Lampkin and Kennedy, 1965; Redfern, 1968; Sacker et al, 1971; Frisch, 1972; Brown and Hutchinson, 1973; Trail and Gregory, 1981). In such situations, the genetic potential for higher productivity of the pure European breed is less important than the superior ability of the zebu to rear a calf without losing too much condition, so that it conceives again.
In the pastoral or subsistence dairy ranching situation it is the relative stability of the small milk yield under fluctuating environmental conditions which is all important. The zebu achieves this stability because it lays down tissue reserves during the rains, whereas high-yielding European cows divert a greater proportion of dietary energy into milk (Trigg and Topps, 1981). As a result the European crossbred cow fails to provide a reliable milk supply in the dry season, because of lower tissue reserves as well as higher metabolic demands. Futhermore in European breeds of cattle, milk yield is depressed by water restriction (e.g. Little et al, 1976), whereas the admittedly smaller milk yield of zebu cattle is largely unaffected by alternate day watering by the time range conditions demand it. The conclusion is that if crossbred cattle produce more milk during the rains, when there is a surplus anyway, and less during the dry season when it is really needed, then they should not be considered an improvement, even if their overall lactation yield is higher.
In Kenya Maasailand, there is a trend with increasing group ranch development towards using introduced zebu bulls, notably the 'improved' milking Sahiwal and 'improved' beef Boran, on the Small East African Zebu. It also happens that the most developed group ranches have the highest rainfall, and it will be interesting to see how far down the aridity gradient the practice of crossbreeding is taken before it becomes counter productive (King et al, in press).
Both the Sahiwal and Boran are bigger than the Small East African Zebu, as well as being more productive. But when forage is scarce the small animal may prove to be the better milker, because it will walk and feed at much the same rate as the larger ones but have a smaller maintenance requirement. Livestock appear to get smaller as the average THI increases (Table 21). The phenomenon appears to apply to wild as well as domestic African ungulates (Dorst and Dandelot, 1970). While poor nutrition, high parasite loads and disease challenge may be contributory factors, dwarfism may be the physiological response to a humid, hot environment. Having accepted the general principle of using small indigenous animals because of their adaptation to a hostile environment, it is inconsistent to still select for bigger size.
The main objection to the use of 'improved' livestock in pastoral areas is their inability to realize their genetic potential in the absence of some improvement in management. At present livestock in much of pastoral Africa do not need to have a high production potential, because the low nutritive value of forage, supplied by grasslands and crop byproducts, allows but a limited yield if not supplemented with arable crops, which is unlikely (Jarrige, 1980).
It has been suggested that the amount of exotic blood in livestock should decline, as the productivity of the pastoral system declines, but the suitability for pastoralism of the genetic material in 'improved' breeds has so far not been questioned. Under conditions of heat, water or nutritional stress the exotic genes may be working at cross purposes with the objectives of the system.
It has also been suggested that pastoral cattle should remain reasonably independent of water. But emphasis on water conservation efficiency means that the criteria evolved in the developed tropics for livestock selection cannot be strictly applied. In the southern United States and northern Australia, research has been concerned with identifying livestock, particularly cattle, which exhibit the least rise in rectal temperature in response to heat stress. This attribute is described as heat tolerance, and has been related to higher growth rates, lower embryo mortality and higher birth weights (McDowell, 1972; H.G. Turner, unpublished). Heat tolerance is primarily due to efficient heat dissipatory mechanisms, notably sweating, but it could also be due to a depression in endogenous heat production which is why heat tolerance tests are not particularly sensitive indicators of productivity (Branton et al, 1966). Lower heat production implies a lower metabolic rate and consequently lower growth potential. The basis for selection has therefore been refined to that of maintaining a normal rectal temperature without a reduction in food intake (Vercoe, 1976; Frisch and Vercoe, 1977; 1978).
If sweating is to be kept at a minimum in the pastoral cow then either high heat loads must be avoided and dry heat loss increased, or the body temperature must be allowed to fluctuate (Figure 8 and section 4.2).
The value of a light or medium coloured, dense coat to reduce solar radiation reaching the skin surface is generally recognized, as is the disadvantage of a 'woolly' coat in cattle which has been negatively correlated with weight gain and milk production in the tropics, probably because it reflects a physiological imbalance (McDowell, 1972).
The best shape for reducing radiant heat load is that of the camel, with its long legs, short, slab-sided body and razor-back. The sale of immature steers may be an important component of production and the objectives of beef production may be in conflict with those for water conservation. For example, the Boran beef breeder is looking for a long, broad, straight back, avoiding rangy animals because they are indicative of slow maturity and stocky ones because they lack the more extensive and expensive back joints.
There is some debate about the efficacy of beef breeding selection programmes in changing the shape of the animal. For example, McDowell (1972) noted that the effect of zebu genes in a crossbreeding programme in the USA was to shorten the length of the back. In a crossbreeding programme in western Uganda, Trail et al (1971) found that at the same weight, there was no difference in side and leg length between Boran, zebu and Redpoll crosses. Ankole crossbreds, which are a Sanga type, were both longer and taller. The only crossbred that was longer backed and shorter legged was the Aberdeen Angus crossbred, reflecting the more intensive selection for blockiness which has gone on in this breed. The equable climate in western Uganda did not seem to favour one crossbred more than another. More extreme climates might have done so, and also altered the shape an-d size of the growing animals (section 4.2.5).
The role of appendages has not been completely clarified and there is not enough evidence to promote them at the expense of other characteristics. Thus skin folds should not extend into pendulous sheaths, nor should horns be retained just for their possible thermoregulatory function. On the other hand there may be good reasons not to dehorn animals in a pastoral situation, where horns may be highly prized. For example the size, shape and colour of the horns of the Ankole cow, which may be up to 85-90 cm long and weigh 7 kg or 1.7% of bodyweight, are second in importance only to coat colour in the eyes of their breeders, the Abahima people (Mackintosh, 1938). Horn disbudding had no significant effect on growth rates from birth to 3 years (Trail and Sacker, 1966), and it is possible that horns would act as radiators in hotter, drier environments.
There are also other behavioural responses to be encouraged, apart from shade-seeking. Camels are adept at exposing the minimum surface area to solar radiation under conditions of water deprivation. When the herdsman notices that all the camels have turned to face the sun, it is time to take them to water (C.R. Field, unpublished). Schmidt-Nielsen (1965) describes how the dehydrated camel sits on the ground with its legs under its body facing the sun, and how groups huddle together couched on the ground to reduce heat flow from the environment during the hottest hours of the day in the Sahara.
The alternative of allowing the body temperature to fluctuate was addressed by Thigh (1972) who suggested that the benefits which thermolability bestows on the camel could apply equally well to cattle, sheep and goats. As has already been indicated (section 4.2.6), smallstock probably do not have a large enough mass to store a significant portion of the day's radiant heat load to make thermolability worthwhile. Cattle are big enough and at least one breed, the N'Dama, allows its body temperature to fluctuate. The mechanism has been associated with trypano-tolerance, but it has also been noted that the N'Dama has low water requirements. Furthermore thermolability does not seem to have affected the N'Dama's productivity compared with other indigenous breeds of cattle (Table 45). Selection for thermolability runs contrary to the philosophy behind the heat tolerance tests mentioned earlier in this section.
Because of the relatively small milk yield of the pastoral cow, it may be tempting to replace it with a beef animal. But the criteria for selecting a pastoral cow are slightly different from those for the beef cow, which may be selected on the basis of her calving interval, ability to maintain bodyweight while suckling a calf, and on the calf's viability and weight at weaning. Admittedly the 'improved' Boran is only a few generations away from its role as a pastoral cow, and may retain the pastoral characteristics. To support this last suggestion it has been shown that 'beef' Brahman cattle imported into tropical Australia, milked more than Herefords and used feedstuffs and nutrients from body stores to lactate at the expense of reproduction (Turner, 1980).
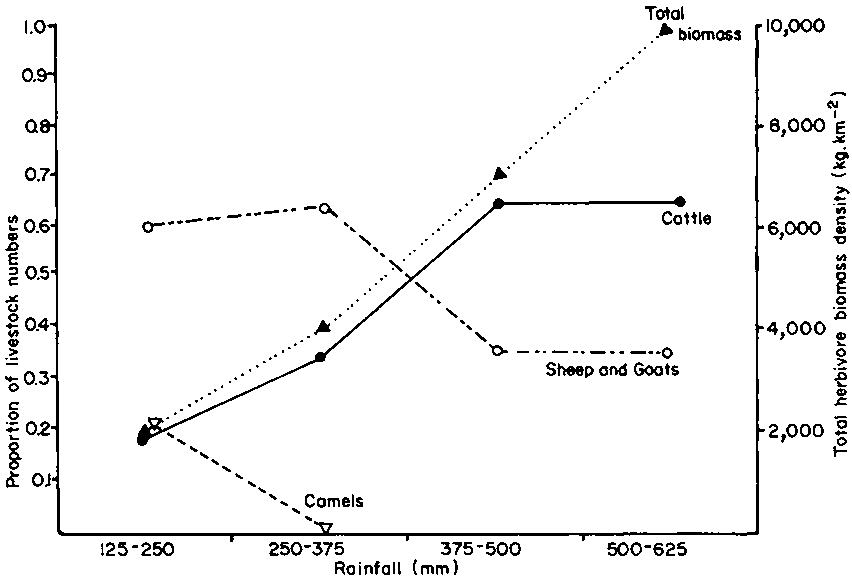
Despite the attributes of the pastoral cow discussed above, pastoralists cannot rely on cattle milk throughout the dry season, and in arid areas they may not be able to keep cattle at all. Thus a mix of species is kept, related to the rainfall in the area (Figure 24). This mix is also affected by such factors as sedentarisation, range trends and exposure to new markets (King et al, in press).
The ratio between the species varies with the rainfall since it is partly related to heat tolerance and water needs. Attributes in water conservation efficiency are normally thought to be traded off against productivity, but enough evidence has been produced in this report to show that this relationship is not constant. In a direct comparison of the water cost of production between species on Galana ranch, it was found that the growth rate of domestic oryx, eland and cattle was of the same order, but that oryx had about one third of the water requirements of the other two (Table 43).
No similar work has been done in the pastoral situation, but one can make a preliminary assessment of whether or not big discrepancies in the water cost of production are likely to occur. The first step is to obtain a measure of the productivity of different species on natural rangeland.
A useful index was developed by ILCA (1979a) which measured productivity in terms of the weight of the yearling calf and the calf equivalent of milk taken by the herdsmen, divided by the weight of the dam maintained annually. The same sort of index was applied to sheep and goats, with the difference that the weight of the progeny was taken at weaning (5 months), although production was still expressed on an annual basis. In order to compare large and small stock in the same index, production has been expressed on an annual basis in terms of what the female weans. Weaning age is taken as 12 months in camels (Bremaud, 1979), 9 months in cattle and 5 months in smallstock. Thus yearling calf weights given in ILCA (1979a) have been corrected to weight at 9 months using the following equation:
9-month weight = 12-month weight - (12-month weight - birth weight)/7 (kg) (6.01)
Table 43. Comparison of daily growtha and water turnover in domestic oryx, eland and Boran cattle.
|
Species |
Water turnover (ml l-0.82) |
Growth |
Water cost of growth (ml.g-1) |
|
Oryx |
68 |
3.2 |
21 |
|
Eland |
202 |
3.1 |
65 |
|
Boran cattle |
188 |
3.4 |
55 |
a From birth to 98% of asymptotic weight (W8).
Source: Carles et al (1981).
Table 44. Annual productivity estimates for the camel.
|
Parameter |
Unit |
Code |
Calculation |
Value |
|
Calving |
% |
A |
|
50 |
|
Calf viability to weaning |
% |
B |
|
60 |
|
Calves weaned |
% |
C |
A × B ÷ 100 |
30 |
|
Calf weaning weight |
kg |
D |
|
150 |
|
Total weight of weaners |
kg % |
E |
C × D |
4500 |
|
Milked out yield |
kg |
F |
|
1460 |
|
Liveweight equivalent of milk |
kg |
G |
F ÷ 9 |
162.2 |
|
Female viability |
% |
H |
|
95 |
|
Females maintained |
% |
I |
H + [(100 - H) ÷ 2] |
97.5 |
|
Female weight |
kg |
J |
|
420 |
|
Female metabolic weight |
kg0.75 |
K |
J0.75 |
92.8 |
|
Per cow maintained |
||||
|
Weight of weaner |
kg |
L |
E ÷ I |
46.2 |
|
Liveweight equivalent of milk |
kg |
M |
G × (A ÷ 100) ÷ 1 |
83.2 |
|
Productivity indexa |
kg |
N |
L+M |
129.4 |
|
Productivity indexa |
g.kg-1 |
|
N × 1000 ÷ J |
308.1 |
|
Productivity indexa |
g.kg-0.75 |
|
N × 1000 ÷ K |
1394 |
a Defined as the total weight of weaned calf plus liveweight equivalent of milk produced per annum.
Source: Mason and Maule (1960); Bremaud (1969); Spencer (1973); Dahl and Hjort (1976); Williamson and Payne (1978); and Wilson (1978a).
Camel productivity traits have had to be derived from a variety of sources (Table 44). The low calf viability to weaning (60%) is a reflection of the demands made by the household. These needs can hardly be reduced: in times of drought the Rendille of northern Kenya let the male camel calves die first, then their own children, and then the female calves (H.J. Schwartz, unpublished).
The values of the productivity index for species and breeds from all over sub-Saharan Africa have been brought together in Table 45.
When the productivity index is expressed in Table 45 as g of weaner produced per kg of dam maintained per year, it can be seen that goats are more productive than sheep, and sheep more than cattle and camels. However, when size is discounted, there is no difference between the 20 kg West African Dwarf goat and the 420 kg camel. It is difficult to know if differences between breeds within sheep or goats are real or due to a paucity of data, compared to cattle. The mean values for sheep, goats, cattle and camels are remarkably similar with a range of 1085 to 1395 g.kg-1W0.75. year-1 (30%). In contrast the values for mean daily water turnover are remarkably different, with a twofold difference between the Small East African goat (at 300 ml.l-0.82) and camel (at 150 ml.l-0.82) in the same environment (Galana).
The conclusion is that differences in the water cost of production between species, if not breeds within species, are worth more study, particularly in the context of pastoral water development.
This management practice involves herding livestock as far from water as possible at the start of the dry season, when the vegetation is green and the days are cool, and gradually bringing them closer as the vegetation dries out and the days become hotter. It is commonly practiced with camels and smallstock, for example by the Rendille and Gabra of northern Kenya (IPAL, 1982), but not with cattle. Herders of cattle normally adopt a practice of centrifugal watering, which involves grazing near water first and extending their range as the dry season progresses. The reason for the different practices probably relates to the lower water requirements of camels and smallstock and their higher water intake from forage, compared to cattle.
Nevertheless, the observation of cattle in Niger suffering fatigue and weight loss but walking maximum distances at the end of the dry season and at the hottest time of year, prompted Rippstein and Peyre de Fabregues (1972) to recommend centripetal watering. The practice was adopted by Klein (1981) in an experiment to determine the optimum stocking rate on Sahelian pastures in Niger. A traditionally managed herd was included as a control, but the comparison of the two systems of watering was confounded by the fact that the animals on the centripetal system were in paddocks, and may even have grazed at night.
The value of centripetal watering in maintaining bodyweight at the end of the dry season may be negated in the pastoral system if it depresses lactation at the beginning. The milk yield will be at its peak at the end of the rains when the cow will be expected to walk as far away from water as possible. An energy budget for that period suggests that the animal should not walk more than about 10 km.d-1 (Table 46).
Table 45. Comparison of the productivity index in different breeds of livestock under subhumid to semi-aria ranching conditions, and low to no tsetse fly challenge.
|
|
|
Weaner per dam per year |
|
|
Species |
|
(g kg-1) |
(g kg-1W0.75) |
|
CAMEL |
|
308 |
1395 |
|
CATTLE |
|
|
|
|
Humpless: |
N'Dama |
325 |
1310 |
|
|
West African Shorthorn |
334 |
1215 |
|
Humped: |
Boran |
349 |
1530 |
|
|
Small East African Zebu |
319 |
1295 |
|
|
Sokoto Gudali |
274 |
1175 |
|
|
White Fulani |
394 |
1690 |
|
Humpless × humped: |
Africander |
256 |
1180 |
|
|
Ankole |
284 |
1230 |
|
|
Tswana |
338 |
1540 |
|
Species mean: |
|
316 |
1375 |
|
GOATS |
|
|
|
|
Dwarf: |
West African |
648 |
1390 |
|
Intermediate: |
Small East African |
428 |
975 |
|
Long-legged: |
Sahel |
363 |
840 |
|
|
Sudan Desert |
618 |
1495 |
|
Species mean: |
|
514 |
1175 |
|
SHEEP |
|
|
|
|
Thin-tailed: |
West African Dwarf |
639 |
1405 |
|
|
Sahel |
434 |
1095 |
|
|
Sudan Desert |
530 |
1335 |
|
Fat-tailed: |
Maasai |
297 |
710 |
|
|
Tswana |
370 |
890 |
|
Species mean: |
|
454 |
1085 |
Source: Adapted from ILCA (1979a) and Table 44 for the camel.
Table 46. Estimated daily energy budget of a zebu cow on centripetal watering.
|
Item |
Absolute value |
Coefficient |
Equation number |
Metabolisable energy (MJ) |
|
Forage intake |
6.34 kg DM |
d = 0.50 |
(5.02) |
46.3 |
|
Maintenance at rest |
250 kg |
|
(5.03/4) |
29.0 |
|
Extra costs: feeding |
46.3 MJ ME |
40 kJ.MJ-1ME |
|
1.9 |
|
lactating |
2.0 kg |
3.6MJ.kg-1 |
(5.05) |
12.9 |
|
walking |
9.5 km |
1.8kJ.kg-1.km-1 |
|
4.3 |
|
Total expenditure |
|
|
|
48.1 |
|
Energy balance |
|
|
|
- 1.8 |
The estimated energy deficit of 1.8 MJ ME. d-1, or 0.4 kg weight loss per week, is unlikely to depress lactation. However the boma can only be located a maximum of 5 km from the watering point. Therefore the cow must be watered every second day so that it can graze away from water on alternate days, thereby increasing the radius of the grazing area from the watering point to about 9 km. The problem with the implementation of 2-day watering when the cow's water needs are still high (due to high DMI and milk yield) is that the animal may become severely dehydrated. This will become apparent if it drinks more than 50 l every second day, i.e. it has a daily drinking requirement of 25 l. The degree of dehydration represented by 50 l every second day in a 250 kg cow is 20%. Any further water needed to offset the demands of DMI and milk yield must come from water in forage, which can be calculated using equation (5.21):
WF = (I × Iw) + (WK × WKw) + (MY
× MYw) - WD
= (6.34 × 4.5) + (0 × 0.35) + (2 × 0.87) - 25
= 5.3 l.d-1
The moisture content of the grass which will achieve this water intake is obtained from equation (5.20):
Forage moisture = WF/(I + WF)
= 5.3/(6.34 + 5.3)
= 0.46
The conclusion reached is that lactating zebu are unlikely to maintain full productivity on a centripetal watering system when the grass moisture falls below 0.46. This value is based on a mass of assumptions which need to be validated. However, the example illustrates the principles involved and one of the main constraints to the innovation.
The other major constraint in Niger, and of course elsewhere, is how to destock around the pumping stations so that centripetal watering can be adopted. The pasture within a radius of 8 km of these pumps is about 20 000 ha and should support <3000 TLU, whereas in practice it carries >10 000 TLU (Rippstein and Peyre de Fabregues, 1972).
When the rains have failed and it is likely to be a difficult dry season the transition to 2- and 3-day watering should be made earlier in order to precipitate a 'siege' condition in the animals. The value of reducing cattle maintenance requirements to two thirds of normal, is that it allows the grazing to last one third longer or carry one third more animals. There is also a saving in tissue reserves compared with an animal on a daily watering regime, which could be in the region of 95 MJ ME (Figure 18). This saving does not look much compared with a daily maintenance requirement of 23 MJ ME until one realizes that the cow with a depressed metabolic rate is approximately in energy balance (Table 31). The decision to change from a production to a survival strategy is normally associated with lack of grazing, but it should also be taken when the quality of the forage is low. As Rogerson (1963) showed and has been mentioned in section 4.3.2, cattle on a roughage diet with a low protein content are better able to maintain energy balance if water intake is reduced.
The trend of decreasing frequency of watering is usually associated with increasing distance of the boma from the watering point, which puts added strain on the water supply to the household. This constraint can be overcome with the provision of baggage animals. In the semi-arid areas donkeys appear to be in plentiful supply, but in the deserts where camels are used there is not enough slack in the system to rear an adequate number of males (section 5.2.2). Consequently there may be a case for providing mature baggage camels to poor settlements to allow them to get away from water into better browsing areas, thereby breaking the vicious circle in which they have neither milk nor forage enough to rear their own males. In feet this is one of the recommendations of the resource management plan for the Rendille area of northern Kenya (IPAL, 1982).
Another category dependent on carried water are young, unweaned livestock in the hottest pastoral environments. For example, in the northern Sahel, the Tuareg keep their camel calves in camp for the first 4 months and give them 5 to 10 l water from a waterskin every 3 to 4 days, and goat kids are kept in camp for 6 months and drink 1 to 21 every day (Swift, 1979). The provision of inadequate water to compensate for the milk taken by man could be one more factor contributing to the high mortality rates in young stock.
Except where intense sun forces herdsmen to let their livestock out before dawn, night grazing is not practiced in pastoral Africa. The main reason is that it is very labour intensive, and also dangerous for the stock (predators) and the herdsmen (snakes). A prerequisite for night grazing is fencing and predator control, both human and animal. The value of night grazing is that it provides extra time to eat vegetation with a higher moisture content at low environmental heat loads. It is practiced by many wild herbivores in desert areas, and makes them largely independent of drinking water (section 3.3.3). In cattle, its most important contribution is to the high yielding dairy cow which can be identified by the extent of its foraging at night (Stobbs, 1975). Good night paddocks are desirable for optimum dairy production on tropical pastures. At the other end of the scale, it was found in Tanganyika that Small East African Zebus allowed to graze at night were only 3 kg heavier after 72 weeks than their counterparts which were penned at night (Meyn, 1970). In a similar comparison, Boran zebus put on an extra 23 kg with night grazing. It was concluded that a small breed of zebu can satisfy its nutritional requirements under traditional pastoral management whereas a larger breed, like the Boran, cannot. In another trial in Tanganyika with Boran and Angus × Boran steers, the 5 to 6% improvement in liveweight of 24 h grazing over night penning was offset by the loss of 2 steers killed by lions (Wigg and Owen, 1973). The same authors noted one benefit of night grazing, not previously mentioned, namely the manure the pasture receives which would otherwise accumulate in the night enclosure. This extra fertilizer visibly assisted the spread of a most useful perennial star grass (Cynodon dactylon).
From the foregoing, it can be seen that the value of night grazing to small breeds of indigenous livestock is not automatic; it also varies with environmental and pasture conditions. For example, when West African Shorthorn cattle were left out during the whole 24 h in Ghana, the amount of time they chose to spend grazing at night was found to vary with the environmental conditions (Rose Innes, 1963). In Uganda, Joblin (1960) found that the restriction of night grazing led to a significant decline of 30% in liveweight gain in zebu oxen, but the difference was largely attributed to periods of moderate grass shortage, when those animals with the longer grazing period were able to select a diet of superior quality and quantity. Under good or very bad grazing conditions the provision of night grazing made no difference. In many areas of Maasailand the nights can be clear, cold and windy, particularly towards the end of the dry season. Crowding cattle together in a thorn boma for the night must reduce their heat loss and their need to waste limited body energy reserves on thermogenesis.
![]()
![]()
![]()
{kind=link}