![]()
![]()
![]()
The purpose of this chapter is to relate the physiological aspects of water and energy turnover discussed in chapters 2 to 4 to the real-life situation as influenced by climate and livestock management. Emphasis will be placed on the dry season when water is scarce, and on the more important livestock species.
The effects of the different seasons on domestic herbivores are well illustrated in a traditional pastoral system because there is little opportunity to alleviate adverse conditions with supplements of food and water.
5.1.1 Energy units
5.1.2 Forage intake
5.1.3 Energy use
5.1.4 Energy budgets
5.1.5 Tissue mobilisation
5.1.6 Response to chronic energy deficit
5.1.7 Compensatory growth
5.1.8 Body water turnover
The way in which the energy use of the grazing animals is affected by different watering regimes can be simulated in simple tables. The level of detail is much coarser than that shown in Figure 19 and described in chapters 2, 3 and 4, but it is beyond the scope of this report to try and develop a complicated model. Instead, a number of assumptions have been made: the environmental heat load has been kept constant between watering regimes, and is assumed to be high. As a result, drinking water intake has been elevated by a constant related to temperature and varied primarily by DMI. The prediction of forage intake has not been discussed so far, and it must be established along with energy to allow the completion of simple energy budgets.
The zebu has been used in this illustration because it is the most important species of livestock in sub-Saharan Africa.
Metabolism in the ruminant is currently expressed in one of two ways derived from different systems of analysis: one based on metabolisable energy (ME), proposed by Blaxter (1962), adopted by the UK Agricultural Research Council (ARC, 1965) and put into practice by the Ministry of Agriculture, Fisheries and Food (MAFF, 1975); the other, based on net energy (NE), proposed by Lofgreen and Garrett (1968) and adopted by the USA National Research Council (NRC, 1970). The relative merits of each system have been reviewed by Webster (1978), who points out that it does not matter which convention one uses provided that one recognizes that it is not possible to combine estimates of the energy value of foods from one system with estimates of the energy requirements of the other and get a sensible answer. The reason is that the expressions are based on different conventions.
For the purposes of the present analysis, ME units have been chosen. Therefore other components of energy metabolism in the ruminant, which is illustrated in Figure 20, must be converted into ME, using coefficients which will be described in the text. Most of these coefficients have been taken from Konandreas and Anderson (1982), and modifications from other sources have been cited.
Forage availability varies with season for a given set of soil and vegetation types and conditions. These seasons have different climatic characteristics in different parts of sub-Saharan Africa: in East Africa there are two dry seasons, one hot and one cool, each year; in West Africa there is only one dry season, which begins cool and becomes hot; and in central and southern Africa there is a single cool dry season Such differences affect water turnover more than energy intake and use. Therefore, one can generalise by stating that at the start of the dry season conditions are uniformly good for food intake; there is usually an adequate crop of standing hay, and intake of nutrients is not depressed by lack of drinking water, by a high parasite challenge or by high THI. Thereafter there is increasing dessication of the vegetation and drying up of watering places as the dry season progresses. Thus there is a steady decline in the quality and quantity of herbage on offer, except where pastoralists exploit floodplain grazing, such as that of the inland delta of the Niger in Mali (Wilson et al, 1983).
Figure 20. Energy metabolism in the ruminant. (Source: A.J.F. Webster and J. M. King (unpublished)).
Within the animal, the process of conversion of feed sources of energy into animal products can be divided into the absolute capacity of the animal to ingest metabolisable energy, and the partial efficiencies of the conversion processes themselves. These are equally important (Blaxter, 1969b).
Before describing the way in which forage intake is currently predicted, it is worth mentioning the reservations that have been expressed about the approach, particularly in the tropical situation. These reservations have been summarised by Freer (1981) as follows: 'There are at least three areas where the simple model based on the amount of herbage and its digestibility avoids important complexities of the real situation.... Firstly it is probable that weight per unit area is an insufficient predictor of the availability of herbage to a grazing animal. Secondly it is certain that digestibility is an inadequate index of those quality changes in herbage wich affect its rate of disappearance from the gut. Van Soest (1982) found that there was a higher correlation between intake and plant cell wall content (- 0.76) than intake and digestibility (0.44) in 187 forage species tested. Freer (1981) continues: And lastly, little is known on how the two variables (weight and digestibility) interact with increases in energy demand, during growth and lactation for example, to increase intake. But he goes on to point out that more experimental information is needed before substantial improvements can be achieved.
In the meantime the amount and digestibility of the herbage on offer will continue to be the principal components of the model of forage intake, particularly as they have been studied comprehensively in an African situation (Elliot and Fokkema, 1961; Elliot et al, 1961). The general relationship obtained to determine intake has been developed by Konandreas and Anderson (1982) to give the following equation:
I = m(d,t). m(Q). m(D). m(t). m(x,t)
m (p,t).a.We-0.73 (5.01)
Each variable is listed below with its range of values likely to be obtained for pastoral cattle in the dry season:
I = dry matter forage intake (kg.d-1)
m(d,t) = digestibility-of-forage-correction multiplier = 1/(1-d), where d = digestibility between 0.40 and 0.65
m (Q) = quantity - of - forage - correction multiplier (t.ha-1), with forage on offer (Q) varying from 0.7 - 2.1 with threshold for intake reduction (Qx) 0.8. Thus
m(Q) = 1 at all values for Q>0.8, and Q/Qx below 0.8
m(D) = grazing-time-correction multiplier, a function of the distance walked per day (D), which above 14 km (D*) reduces forage intake by 1.0 - 0.05 (D-D*); otherwise m(D) = 1.0
m(t) = age-correction multiplier = 1
m(x,t) = saxcorrection multiplier = 1
m(p,t) = physiological-status-correction multiplier, which for dry females = 1, and for lactating cows = 1.15, partially due to an increase in chewing and ruminating (Dulphy et al, 1980)
a = intake coefficient (kg DM.kg-0.75.d-1) which is equivalent to the rate of pas sage, and taken as 0.042 for dry cows and 0.049 for lactating cows
We = expected liveweight (kg) for animal's age and sex.
The gross energy content of tropical pasture is relatively constant (17.2 to 18.7 MJ.kg-1DM), giving a mean value of about 18 MJ.kg-1, which is about 0.5 MJ.kg-1 less than the mean for temperate pastures (Minson, 1981). As shown in Figure 20, portions of this energy are lost as faeces, urine, methane, and fermentation heat during the conversion to ME. Thus the energy (E) from a given intake (I) can be expressed as:
E = I × 18 × d × 0.81 MJ ME (5.02)
Energy demands vary with the body functions for which energy is used. Thus the net energy (NE) demands of the product (e.g. lactation, growth) are converted back to ME using different coefficients. For maintenance the NE, or total metabolic heat production, is estimated from a specific variant (Table 24) of the general formula for fasting metabolism: 0.293 Wt kg0.75 (MJ.d-1), more familiar in its original form of 70 Wt kg075kcal.d-1 (Brody, 1945; Kleiber, 1947). The amount of ME used for maintenance in the zebu cow can be estimated from the equation:
Em = 0.343 Wt0.73/km MJ ME (5.03)
where km is the efficiency with which ME is used for maintenance and
km = 0.55 + (0.3 × 0.81 × d) (5.04)
To maintain bodyweight, the cow must also graze and walk to water, which increases energy expenditure above basal rates by 30-70% (Young and Corbett, 1972; Lewis, 1975).
The cost of prehension, tearing and eating coarse dry grass has been estimated at 40 kJ.MJ-1 ME of ingested food, based on Webster (1980).
The energy cost of walking per horizontal km is similar for Bos taurus and Bos indicus cattle on a diet at or above maintenance. The most efficient speed is between 2.5 and 3.5 km. h-1 when the cost is 1.8 kJ.kg-1 (Ribeiro et al, 1977; Finch and King, 1979). At faster and slower speeds the energy cost per metre increases.
Productive uses of energy include growth, pregnancy and lactation. The ME used for lactation is calculated from the NE required for milk, divided by the efficiency with which ME is conversed to NE for lactation. This efficiency (k1) is less than for maintenance:
k1 = 0.463 + (0.24 × 0.81 × d) (5.05)
The NE requirements for milk are calculated from the milk yield (kg.d-1) multiplied by its energy content (e1). The energy content is given approximately by the following equation:
e1 = 0.0386BF + 0.205SNF - 0.236 (5.06)
where
BF = butter fat content (g.kg-1), and
SNF = solids-not-fat content (g.kg-1)
For the zebu, BF = 54 g.kg-1 and SNF = 85 g.kg-1 (Williamson and Payne, 19783 while e1 = 3.6 MJ.kg-1.
The efficiency with which surplus ME is converted into energy for weight gain (kg) is lower than that for lactation. It is obtained by multiplying ME by kg where:
kg = 0.03 + (0.81 × 0.81 × d) (5.07)
However liveweight gain during lactation is achieved at a similar level of efficiency as lactation, thus:
kg = k1 (5.08)
The liveweight gain from the available NE derived from equation (5.07) or (5.08) is calculated as follows:
DW = Eg/(6.28+0.018W+0.3Eg) (5.09)
where
DW = liveweight gain (kg.d-1)
Eg = available NE (MJ.d-1)
W = present liveweight (kg).
Table 24. Fasting metabolism of livestock.
|
Species |
Weight (Wt) |
Fasting metabolism |
Equation |
Source |
|||
|
Predicted |
Measured |
||||||
|
Donkey |
160 |
13.2 |
9.6 |
0.73 (0.289Wt0.75) |
CEEMAT(1972) Robinson and Slade (1974) BDPA (1974) |
||
|
Horse |
350 |
23.7 |
24.3 |
0.289Wt0.75 |
CEEMAT(1972) Robinson and Slade (1974) BDPA (1974) |
||
|
Bovine |
|||||||
|
|
humped |
350 |
23.7 |
24.7 |
0.343Wt0.73 |
Rogerson (1968) |
|
|
|
humpless |
|
|
|
|
|
|
|
|
|
N'Dama |
260 |
19.0 |
- |
- |
|
|
|
|
European |
500 |
31.0 |
35.2 |
0.376Wt0.73 |
Blaxter (1969a) |
|
|
|
European |
600 |
35.5 |
37.0 |
0.305Wt0.75 |
Flatt et al (1969) |
|
Camel |
450 |
28.6 |
20.4 |
0.209Wt0.75 |
Schmidt-Nielsen et al (1967) |
||
|
Eland |
300 |
21.1 |
30.2 |
0.464Wt0.73 |
Rogerson (1968) |
||
|
Goat |
30 |
3.8 |
3.2 |
0.251Wt0.75 |
Macfarlane and Howard (1972) |
||
|
Sheep |
|||||||
|
British |
50 |
5.5 |
4.2 |
0.243Wt0.73 |
Blaxter (1969a) |
||
a Source: Kleiber (1947; 1969).
Simple budgets can now be compiled to simulate real life situations, which are developed further in section 5.2.1. First the parameters listed in Table 25 are derived, in this case from sample data given by Konandreas and Anderson (1982).
The factors affecting energy intake and output can now be estimated separately. For example, forage intake (I) is calculated by inserting the values from Table 25 into equation (5.01):
I = 1/1 - 0 50.1.0 × 8.1.1.1 × 15.0 × 049.2500.73
which yields an intake of 5.08 kg DM.d-1. This value is entered into the energy balance table (Table 26), where it is expressed as MJ ME, using equation (5.02).
It can be seen from the example given (Table 26), that feed intake is insufficient to meet the animal's total energy requirements. The deficit consists of 12.9 MJ ME for lactation and 0.7 MJ ME for maintenance, and is made good by mobilisation of body tissue. The energy value of the tissue mobilised is taken as 20 MJ.kg-1 (MAFF, 1975), although the mean value for the meat in the carcass may be lower (Ledger and Sayers, 1977). It is used with an efficiency of 0.84 to produce milk energy (NE) (ARC, 1980).
The energy yield of milk from each kg of body tissue mobilised is:
20 × 0.84 = 16.8 MJ NE (5.10)
This is equated to dietary ME by dividing by k1 (equation 5.05):
16.8 ÷ 0.560 = 30 MJ ME
The amount of tissue mobilised for milk production is therefore:
12.9 ÷ 30 = 0.43 kg
The same efficiency coefficient is assumed when energy released from body tissue mobilisation is used to meet maintenance requirements (Konandreas and Anderson, 1982). Therefore the amount of tissue mobilised for maintenance is:
0.7 ÷ 30 = 0.02
Thus, total daily tissue loss is 0.45 kg.
Table 26 illustrates the 'typical' energy status of a zebu cow about one third of the way through the dry season. Thereafter the quality and quantity of herbage will decrease still further, so that the estimated daily bodyweight loss will increase above 0.45 kg and one might expect the animal to be dead within 2 months. The fact that it is not can be explained by a number of changes that occur in its metabolism, and in its management by the herdsman. These changes must be quantified where possible.
Table 25. Basis for estimation of daily energy budget of a zebu cow watered daily.
|
Parameter |
Absolute value |
Unit |
Functional form and value | |
|
Forage digestibility |
0.50 |
fraction |
d |
= 0.50 |
|
Herbage (DM) on offer |
2.10 |
t.ha-1 |
m(Q) |
= 1.00 |
|
Distance walked |
18 |
km |
m(D) |
= 0.80 |
|
Milk yield |
2 |
kg |
m(p,t) |
= 1.15 |
|
Intake coefficient |
0.049 |
fraction |
a |
= 0.049 |
|
Expected liveweight |
250 |
kg |
We |
= 250 |
Source: Konandreas and Anderson (1982).
Table 26. Estimated daily energy of a zebu cow, based on Table 25.
|
Activity |
Absolute value |
Coefficient |
Equation number |
ME (MJ) | |
|
Forage intake |
5.08 kg DM |
|
(5.02) |
37.0 | |
|
Maintenance at rest |
250 kg |
|
(5.03/4) |
28.8 | |
|
Extra costs: |
|
|
|
| |
|
|
feeding |
37.0 MJ ME |
40kJ MJ-1 ME |
|
1.5 |
|
|
walking |
18 km |
1.8kJ.kg-1.km-1 |
|
8.1 |
|
|
lactation |
7.2 MJ NE |
|
(5.05) |
12.9 |
|
|
|
|
Subtotal |
50.6 | |
|
Energy balance |
|
|
|
-13.6 | |
In their study of the response of zebu cattle to protracted periods of undernourishment caused by reduced intake of a high quality diet, Ledger and Sayers (1977) concluded that the 33% drop in maintenance requirements was partly due to increased conversion efficiency. But Finch and King (1979) found that the reduction in fasting metabolic rate of zebu steers, in response to a progressive reduction in food, was one third, thus accounting for all the difference observed by Ledger and Sayers. The drop in maintenance demands during the dry season will therefore be quantified using metabolic rate only (Figure 21).
Further conservation of energy is achieved when the cow depresses its milk yield, and the herdsman decreases the distance walked by the animal per day by reducing the frequency of watering. Both these variables have already been quantified in section 5.1.3, but in the case of the undernourished animal there is a significant (P <0.01) reduction in the cost of walking compared with the cow on a maintenance diet. For both classes of animal, the energy cost of horizontal walking, above that of standing still, has been plotted using data from Finch and King (1979) (Figure 22). For 220 kg zebu steers at maintenance the equation is:
Ew = 3.012x - 3.225 (r2 = 0.82)
and for 200 kg steers on a half maintenance ration:
Ew = 2.068x - 2.122 (r2 = 0.81)
where
Ew = energy cost of walking (kJ ME.kg-1.h-1),
andx = speed of walking (km.h-1).
At speed of between 2 and 5 km.h-1 the energetic efficiency (kJ ME.kg-1.km-1) of walking is higher in the undernourished zebu (Table 27). Energy saved by walking more slowly must be balanced against loss of grazing time.
There are two other energy and water saving mechanisms which have not been quantified but which should be mentioned, namely nitrogen recycling and thermolability.
As the diet becomes more fibrous and therefore less digestible, food intake is primarily limited by physical processes. The ruminant stops feeding when a certain level of fill is reached in the reticulo-rumen. Forage will be retained in the rumen until it has been reduced to small particles of mean length 0.5 mm for cattle and 0.25 mm for sheep. The reduction is achieved by microbial degradation and chewing. Low digestibility is correlated with low nitrogen content of the diet, which in turn depresses the cellulytic activity of rumen microorganisms (Dulphy et al, 1980). Konandreas and Anderson (1982) accounted for low digestibility by an additional multiplicative adjustment of voluntary intake at levels of crude protein <6% and digestibility <40%. But lack of crude protein is partly offset by recycling urea from the kidneys to the digestive tract via the bloodstream. This mechanism is present in ungulates employing either the rumen or the large intestine for microbial digestion, and is particularly well developed in species frequently exposed to low protein diets such as the zebu and the camel (Livingston et al, 1962; Schmidt-Nielsen, 1975). This source of non-protein nitrogen is incorporated into microbial protein, thereby increasing the microbial population and its digestion of cellulose before it in turn is digested.
The mechanism of thermolability allows the body temperature to drop below normal at night without increasing thermogenesis, and to rise above normal during the day by regulating sweating rates at a low level (section 4.2.6). The result is body conservation of water and energy. The mechanism of thermolability is initiated in response either to dehydration or to depressed food intake.
The extent to which the mechanisms, described in the previous section, come into play will depend on whether the season is a normal dry one or a drought.
The word drought tends to be used rather loosely. There is a need to distinguish between a dry year and a drought year, as well as drought conditions in a dry year, because the causes and effects can be different.
Drought means dryness. It is a serious deficiency of water for herbage production caused by the cumulative effect of above-average evaporation and/or below-average rainfall (Munn, 1970). In sub-Saharan Africa, rainfall is the more variable of the two components. The level at which low rainfall represents a drought and how it should be calculated is a matter of opinion. In the semi-arid areas of Kenya it has been found that the distribution of values for annual or seasonal rainfall is normal. Instead the probability curve is skewed by the inclusion of occasional very wet years. Thus the value for mean annual rainfall is higher than the median. For the purposes of range and animal management, rainfall expressed as a mean is less useful than median and quartile probabilities (Bille and Heemstra, 1979). It is suggested that a drought year is one in which the total annual rainfall is at or below the lower decile value on the probability curve. The effect of low rainfall on primary production will also depend on the amount of rain in each shower and the interval between showers, bearing in mind that about 25 mm of precipitation is required for growth of perennial grasses and shrubs and 40 to 60 mm for seed germination of annual grassland (J.C. Bille, unpublished).
Table 27. The effect of sub-species and feeding regime in steers on the energetic efficiency of walking.
|
|
|
|
Energetic efficiency (kJ ME. kg-1.km-1) | |||
|
Sub-species |
Weight (kg) |
Feeding level |
2km.h-1 |
3km.h-1 |
4km.h-1 |
5km.h-1 |
|
Bos taurus |
500 |
Maintenance and above |
2.0 |
1.8 |
2.8 |
2.0 |
|
Bos indicus |
220 |
Maintenance |
1.4 |
1.9 |
2.2 |
2.4 |
|
Bos indicus |
190 |
Half maintenance |
1.0 |
1.4 |
1.5 |
1.6 |
Source: Ribiero et al (1977); Finch and King (1979).
The combined effects of sun, wind and feeding by invertebrate herbivores is gradually to remove the leaves from the herbage so that all that remains is a standing crop of stems. Thus the nutritive value of the rangeland will fall below the level for cattle maintenance whether or not it is grazed. On Galana ranch, Kenya, in March 1971, after only 170 mm of rain had fallen over 16 months, the standing crop of Brachiaria spp. had a crude protein content of 3.5%, while Schoenfeldia transiens had 4.5%. These two grasses comprised most of the diet of the grazing stock, which could not supplement their diet with browse because most of the palatable trees and shrubs had lost their leaves. Under such conditions, the response of different herbivores varied: Boran cattle lost so much condition that they had to be moved long before the grazing was exhausted, whereas domestic oryx in the same paddocks continued to grow (King and Heath, 1975). One of the reasons for the difference between the two ruminants appeared to be the ability of the oryx to select what was left of the more nutritious plant parts, whereas cattle with their broad mouths were ingesting a lot of grass stems (Field, 1975). The very fibrous diet of the cattle with inadequate nitrogen content to support normal rumen microbial activity obviously slowed digested passage rate and hence intake. Thus the cattle were starving in paddocks full of grass. Furthermore the provision of water daily, as is the case on a well-run commercial ranch, may have delayed the transition of the animals' metabolism to a 'siege economy'. As a result the body tissue reserves may have been depleted at a faster rate than if they had been forced onto a restricted watering regime earlier in the dry season. Eventually, in this type of situation, the quantity as well as the quality of the forage will become limited.
Drought conditions can also occur in a series of dry, but not drought, years. They occur as a result of overgrazing so that the quantity rather than the quality of the herbage is the main limiting factor. The use of the word 'drought' attributes the blame to the climate and obscures the fact that the conditions often arise because of a lack of range management.
Although the onset of the rains is greeted with relief, it may precipitate many deaths in a starving livestock population. At the end of a drought, animals have practically exhausted their energy reserves. The first heavy rain lowers the ambient temperature and soaks the animal's hair, thereby reducing the insulating effect of its coat. Heat loss from the animal exceeds heat gain and triggers off thermogenesis and shivering, albeit at a lower critical body temperature than in an animal in good condition (Finch and King, 1982). This waste of the last nutritional energy reserves is not immediately compensated for by improved energy intake. In the first place, unless there is a green flush following a fire which proceeded the rain, it takes a few days for the grass to grow after the first shower. Secondly the abrupt transition from sparse, dry grass stems to highly nutritious moist green grass causes the animal to scour (French, 1956b). In fact rainfall over a 21-day period always appears to have a negative effect on liveweight gain (Potter, in press).
For example:
LWG = 442.22 - 9.82a + 9.84b + 0.83c
(r2 = 0.68)
where
LWG = liveweight gain (g/head/d)
a = rainfall within last 3 weeks (mm)
b = rainfall 3 to 6 weeks ago (mm)
c = rainfall 6 to 9 weeks ago (mm).
If the cow survives the dry to wet season transition, it usually starts putting on weight even more rapidly than would be estimated from its ingestion of green grass with a high digestibility (e.g 0.65 in ILCA, 1975). Part of this gain has been attributed to an increase in the weight of the alimentary tract. Gut contents have a water content of 85%, so that one might expect the total body water of the animal to increase from 70% to about 72%, whereas it has been found to drop to 65%, and on carcass analysis it was revealed that fat had been deposited (V.A. Finch and J.M. King, unpublished). An explanation for this phenomenon of compensatory growth is that the demands for maintenance form a much lower proportion of the diet in an emaciated animal than in one in good condition. This is so because the animal has an increased efficiency of food conversion, a smaller proportion of active tissue in the carcass and a depressed metabolic rate. During the weeks it takes to rehabilitate the animal there is therefore a larger surplus of ME available for body tissue deposition than might be expected.
Forage intake during the rains is not a constraint to animal production. Nevertheless there are a number of factors which are counter-productive. For example, the rainy season provides conditions favourable to the rapid multiplication of other forms of life, such as insects and helminths which irritate and sap the productivity of the herbivore. Ambient temperature is not at its highest, but the water vapour pressure of the air is. The combination of the two may produce peak THI readings - for instance at Tahoua, Niger in the Sahel (Figure 11) - and could conceivably cause discomfort even in zebu cattle. Humidity values of 75% r.h. may not be sufficient to reduce the efficiency of evaporative cooling, but the associated cloud cover, which insulates the earth and the animal from radiation to the night sky, may reduce heat loss. If the animal cannot dissipate the large amount of endogenous heat associated with maximum productivity, it will cut back its food intake and metabolic rate, whatever the state of the vegetation.
In order to understand the effect of different management strategies in the dry season, total body water turnover (TBWT) must be divided into the components for which it is used, and also the sources from which it comes. Very little work has been done on livestock under African ranching or pastoral conditions. I have had to rely on my own data on TBWT in zebu steers on Galana ranch in Kenya and extrapolate from there.
The TBWT of steers during the long dry season (June to October), when the mean ambient temperature is about 25ºC, is 150 ml.l-1.d-1 (Figure 9). Given a bodyweight (Wt) of 250 kg and a body pool size of 168 1 (67% Wt), then the absolute volume of TBWT is 25.2 l.d-1.
The two most important costs contributing to TBWT are evaporative cooling of the heat load, and water for forage intake. Unfortunately these two costs have not been separated here, anymore than they have been in the information on farm animals in the British Isles (Table 19). However, a model on heat exchange is being developed (D.M. Swift, unpublished), which should allow separation in future. For the time being, the water cost of forage intake is related to a particular temperature which is assumed to remain constant for each example within a species (section 5.2). The forage intake (I) can be predicted from equation (5.01), assuming in this case a digestibility of 0.50:
I= 1/(1-0.50).1.1.1.1.1.0 × 042.2500.73
I = 4.73 kg.d-1
The energy value of this intake is 34.5 MJ ME according to equation (5.02).
Because the animal is not lactating, nor walking great distances, the TBWT can be related entirely to forage intake at that temperature, assuming the animal is at maintenance. When metabolic water (WM) and respiratory and cutaneous water (WRC) have been deducted from TBWT, the balance represents water from food (WF) and drink (WD) and the values are directly comparable with those in Table 19.
WF + WD = TBWT - (WM + WRC) (5.11)
Metabolic water can be estimated in two ways when energy intake or expenditure is known (section 3.3.5). For example:
WM = 0.029 × total MJ ME (5.12)
WM = 0.029 × 34.5 = 1.00 l.d-1
Alternatively WM can be roughly estimated from TBWT using the following equation from King at al (1978):
y = 0.013k-0.783 (r2= 0.86) (5.13)
where
y = WM as fraction of total input, and
k = flux rate of body water pool (fraction of body pool.d-1).
Respiratory and cutaneous water intake (section 3.3.4) is estimated as follows:
WRC = 0.11TBWT (5.14)
WRC = 0.11 × 25.2 = 2.77 l.d-1
Equation (5.11) can now be solved:
WF + WD = 25.2 - (1 + 2.77) = 21.43 l.d-1
Thus the water cost, in food and drink, of forage intake is:
Iw (WF + WD)/I (5.15)
Iw = 21.43/4.73 = 4.5 l.kg-1DMI
This value for the coefficient Iw falls within the range- for cattle breeds in the British Isles (Table 19) although the environmental temperatures may have been calculated differently in the British and Kenyan situations. A lower value of 2.7 l. kg-1DMI was calculated for zebu cattle in the semi-desert of northern Kenya based on measurements by IPAL (1982) of DMI and water drunk. The herd average for DMI was 2.9% bodyweight compared with 1.9% in the Galana example.
There is an extra water cost of walking (WKW) in high solar heat loads (section 4.1.3):
WKw = 0.0014 × Wt l.km-1 (5.16)
where Wt is the weight of the steer in kg
WKw = 0.0014 × 250 = 0.35 l.km-1
In the hydrated animal at maintenance or above, this water cost is incurred between about 10.00h and 16.30h local time. If the herd leaves from and returns to the night enclosure at 08.00h and 18.30h respectively, then theoretically it can walk for 12 km (4 h at 3 km.h-1) without increasing its water loss. In practice, cattle usually drink between 10.00h and 14.00h, so part of the 12 km (e.g. 6 km) will incur an extra water cost. Thus:
Water for walking = WKw (WK - 6) (5.17)
where WK is distance walked (km).
It should be noted that in undernourished cattle, sweating rates remain high for a much shorter period of the day, namely from 11.00h to 14.00h (Finch and King, 1982). Therefore the water cost of walking must be adjusted to a lower estimate of the distance walked during those 3 hours.
Another extra water cost for the lactating cow is the water secreted in the milk (MYw) which has been estimated from Table 19 as:
MYw = 0.87 l.kg-1MY (5.18)
The sources or inputs to TBWT can be stated as:
TBWT = WF + WD + WM + WRC (5.19)
(The relation of water intake from forage (WF) to forage moisture content is illustrated in Figure 4, and described by the equation:
WF = (I × forage moisture)/(1 - forage moisture) (5.20)
Water drunk can be estimated in a number of ways, by solving equation (5.11) or (5.19) for WD, by combining equations (5.15/16/17/18):
WD = (I × Iw) + (WK × WKw) + (MY × MYw) - WF (5.21)
or by estimating the maximum volume the animal can drink during one visit to the watering point (section 4.3.1) and dividing by the number of days between drinks. The estimation of metabolic water (WM) has already been given in equations (5.12/13) as has WRC (equation 5.14); but where TBWT has not been measured but its components have, WRC can be estimated:
WRC = 0.12 (WF + WD + WM) (5.22)
Once the analytical background has been established it is possible to estimate the water costs and sources of TBWT (Table 28), and use them to construct a table of daily water intake (Table 29). The example chosen is the zebu cow whose energy budget was shown in Table 26.
The values in Table 29 look realistic: the lactating cow is drinking 28 l.d-1 compared with a practical guideline for the dry cow of 25 l.d-1 (Table 10); adjusted body water turnover, by subtraction of metabolic and respiratory water, is 117 ml.kg-1.d-1 compared with a range of 51 to 150 ml.kg-1.d-1 (Table 13); TBWT is 201 ml.l-1 compared with a range of 97 to 274 ml.l-1.d-1 (Table 18).
The maximum values for TBWT were associated with a daily watering regime in the hot dry season. Higher rates, or maintenance of the same rate on an alternate day watering regime, would result in severe dehydration. Therefore body water conservation mechanisms are assumed to become increasingly important at the expense of evaporative cooling which must give way to dry heat loss. The latter mechanism is assisted by a weather pattern of clear nights and substantial diurnal temperature fluctuation, which is from 22 to 35°C in parts of Kenya.
Table 28. Basis for estimation of daily water intake in a zebu cow.
|
Item |
Symbol |
Amount |
Source |
Application to equation |
|
Forage intake |
I |
5.08kg DM |
Table 26 |
(5.20) |
|
Forage moisture |
- |
0.15 fraction |
Table 14 |
(5.20/21) |
|
Water cost of I |
Iw |
4.51.kg-1 DM |
equation(5.15) |
(5.21) |
|
Distance walked |
WK |
18.0 km |
Table 25 |
(5.16/17) |
|
Cow weight |
Wt |
250 kg |
Table 25 |
(5.16) |
|
Water for walking |
- |
4.21 |
equation(5.16/17) |
(5.21) |
|
Milk yield |
MY |
2.0 kg |
Table 25 |
(5.18) |
|
Water in milk |
MYw |
1.741 |
equation(5.18) |
(5.21) |
|
Total ME |
- |
50.6 MJ |
Table 26 |
(5.13) |
|
Body water pool |
TBW |
0.671.kg-1 |
estimate |
- |
Table 29. Estimated daily water intake of a zebu cow.
|
Water source |
Body water turnover | |||
|
Item |
Equation |
l |
ml.kg-1 |
ml.l-1 |
|
Forage |
(5.20) |
0.9 |
4 |
5 |
|
Drink |
(5.21) |
27.9 |
112 |
166 |
|
Metabolic |
(5.12) |
1.5 |
6 |
9 |
|
Respiratory and cutaneous |
(5.22) |
3.6 |
14 |
21 |
|
Total input |
|
33.9 |
136 |
201 |
Source: Table 28.
When a zebu cow stops maintaining a constant body temperature by evaporative cooling, and instead allows its body temperature to rise during the day and fall at night, its feed intake and hence its nutritional energy circuit is likely to be depressed (Figure 19).
The purpose of this section is to show the effect of traditional management practices on water and energy budgets of the most important species of livestock in pastoral areas. Each simulation is preceded by a brief description of the role of each species in the traditional pastoral system. Because some sort of development has already occurred in much of Africa, traditional systems are defined as those not requiring large inputs of skills, equipment or money from outside (Sandford, in press).
The management of cattle for subsistence dairy ranching is different from that for beef. On a commercial beef ranch it may be appropriate to dry off the cow as soon as possible in the dry season, whereas the opposite may hold true in the pastoral situation. The pastoral zebu cow may have to maintain her lactation despite a declining plane of nutrition, decreasing watering frequency, increasing distances between grazing and drinking water, and an increasing environmental heat load. The cow must be able to draw on body reserves for lactation when its nutrition is inadequate, and rebuild these body reserves when grazing improves, rather than increase the overall lactation (Lampkin and Lampkin, 1960b). Seasonal weight loss is also an important means of supporting a higher livestock population, and of achieving a higher productivity per ha, in an arid, water-limited environment than would be possible if bodyweight were maintained (Western, in press). When the rains come there is more than enough forage for the surplus livestock population. For instance, the carrying capacity of an area north of Lake Chad, in Niger, was estimated at 65 000 TLU in the dry season and 200 000 TLU during the rains (Rippstein and Peyre de Fabregues, 1972).
The race chosen for simulation is the Small East African Zebu, indigenous to Mausailand. It is a small cow with a mean adult female weight of only 250 kg (King et al, in press) and a peak daily milk yield of 3.5 kg 4 months post partum (Semenye, 1982). The energy budget and water turnover of this animal have been calculated at three phases of the long dry season associated with daily, 2-day and 3-day watering, assuming the following scenario: the animal calved in March just before the long rains, which were below average, so that its milk yield has been depressed, and by the beginning of July the daily yield is down to 2.0 kg, of which the family are only milking out 0.5 kg.
At this stage, the cow is still drinking every day and grazing on the way to and from the watering point. The boma is located 8 km from water and animal is walking 18 km per day. Up to now, the grazing has been adequate and the cow has lost only 10 kg of bodyweight, but forage will become scarce in the near future.
When it does, the boma will be moved to a second location 11 km from water and the cow watered every second day. On one day the cattle are herded away from water over adjacent peripheral grazing, which is adequate in quantity but has declined in quality. The cow walks about 11 km. The next day the herd is walked in an almost straight line to the watering point and returned home the same evening, covering a distance of 23 km (Western, 1975). The specific effect of 2-day watering reduces milk yield by <5% below the level of daily watering (Semenye, 1982). However the continuing energy deficit and declining plane of nutrition combine to reduce the daily yield to 1 kg.
If the dry season is particularly harsh or extended, or the area severely overstocked, the cattle may be placed on a 3-day watering regime. They then have to search for forage further away from the main boma, because the adjacent peripheral grazing areas have been denuded. Consequently they are unable to return home on the same day, and have to spend the night at a more distant temporary boma, returning to main boma on the second night. The first and second days are thus devoted to grazing, which involves walking about 10 km.d-1.
On the third day the cattle have to get to water by the shortest route, i.e. 11 km, and after they have drunk there is usually only time to take the direct route home. On the third morning, the cattle could be in the desperate straits described by Classen (1977) (see also section 4.1.3) but according to French (1956a) 'this steady dehydration does not lead to unruly behaviour when the animals approach water-holes. In fact, it is a common dry-season sight in Maasailand to observe thousands of cattle waiting patiently in their turn to drink. On reaching the water, they drink steadily for several minutes during which they swell visibly and then stand for 10 to 15 minutes before taking a further short drink prior to the usual 10-mile return journey to their night quarters. Apparently the 3-day watering regime cannot be continued for long, but drinking on alternate days is a system which can be continued for weeks without apparent harmful effects on the animals.'
Three-day watering during the dry season is less common in Maasailand nowadays, because of water development. But it is still practiced elsewhere on the continent, for example in East Africa by the Borana (Helland, 1980) and the Turkana (N. Dyson-Hudson, unpublished), and in West Africa by the Maure (H. Breman, unpublished) and the Tuareg (Swift, 1979).
Assuming the above scenario, the changes in forage availability are listed in Tables 30, and the estimates of energy budget and water turnover in the cow in Table 31. The calculations required to convert the values from one table to the other have been shown in section 5.1, where Tables 25, 26, 28 and 29 give almost identical figures to those listed in the daily watering column of Tables 30 and 31.
Table 30. Basis for calculating daily energy budget and water turnover of a zebu cow on daily, 2-day and 3-day watering.
|
Item |
Unit |
Basis for energy budgets |
||
|
Daily watering |
2-day watering |
3-day watering |
||
|
Forage digestibility |
fraction |
0.50 |
0.45 |
0.40 |
|
Herbage on offer |
tDM.ha-1 |
1.8 |
1.6 |
0.8 |
|
Distance walked |
km |
18 |
17 |
14 |
|
Milk yield |
kg |
2 |
1 |
0 |
|
Intake coefficient |
fraction |
0.049 |
0.049 |
0.042 |
|
Expected liveweight |
kg |
250 |
250 |
250 |
|
Forage intake |
kg DM |
5.08 |
4.90 |
3.94 |
|
Actual liveweight |
kg |
240 |
230 |
215 |
|
Walking cost |
kJ ME.kg-1.km-1 |
1.8 |
1.8 |
1.4 |
|
|
|
Basis for water turnover |
||
|
Forage moisture |
l.kg-1 |
0.15 |
0.12 |
0.10 |
|
Walking in the sun |
km |
12 |
6 |
0 |
|
Body pool |
l.kg-1 |
0.67 |
0.68 |
0.70 |
Source: Tables 25, 26, 28 and 29.
Table 31. Estimated daily energy budget and water intake of a zebu cow on daily, 2-day and 3-day watering.
|
Item |
Energy budget (MJ ME) |
||
|
Daily watering |
2-day watering |
3-day watering |
|
|
Forage intake |
37.0 |
32.1 |
23.0 |
|
Maintenance at rest |
27.9 |
27.6 |
17.9 |
|
Extra costs: feeding |
1.5 |
1.3 |
0.9 |
|
walking |
7.8 |
7.0 |
4.2 |
|
lactation |
12.9 |
6.5 |
0 |
|
Total expenditure |
50.1 |
42.4 |
23.0 |
|
Energy balance |
-13.1 |
-10.3 |
0 |
|
|
Water intake (1) |
||
|
Forage moisture |
0.9 |
0.7 |
0.4 |
|
Drink |
27.7 |
24.2 |
17.3 |
|
Metabolic |
1.5 |
1.2 |
0.7 |
|
Respiratory and cutaneous |
3.6 |
3.1 |
2.2 |
|
Total input |
33.7 |
29.2 |
20.6 |
Source: Tables 25, 26, 28 and 29.
The first column in Table 30 relates to the beginning of July when the cow is still lactating and drinking daily, but at the expense of its own body tissue. The daily energy deficit is 13.1 MJ ME which, according to section 5.1.5, gives a weight loss of 0.45 kg.d-1, 3.15 kg. week-1, and 12.6 kg. month-1. These figures are based on the status of the animal at the beginning of the month and do not take into account the daily decline in weight and milk yield and hence also the decline in energy demands for maintenance, walking and lactation as well as in forage quality, quantity and intake. Nevertheless, it is reasonable to suppose that, by the end of the month, the cow will weigh not more than 230 kg and a change in management will be required.
The change to 2-day watering in August has the effect of increasing forage accessibility by extending the grazing orbit away from the water point, but without increasing the distance walked daily. Nevertheless, there is still a daily energy deficit of 10.3 MJ ME despite the drop in milk yield, which is where the greatest energy saving is made. The deficit represents a weight loss of 2.36 kg per week which may be tolerated for another month. By September the cow weighs only 220 kg and has probably stopped lactating, so that weight loss is reduced to less than 1 kg per week.
As there is no sign of rain in October, the cattle are forced onto a 3-day watering regime because of the scarcity of herbage within the existing grazing orbit. The grazing that is now brought within range is nevertheless of poor quality because it is so late in the season. Consequently, energy intake is depressed to only 60% of the levels given for daily watering 3 months earlier. But the animal is in energy balance for the first time in about 4 months. The reason is that the chronic energy deficit coupled with intermittent partial dehydration has stimulated the conservation responses described in section 5.1.6.
The daily body water turnover follows a similar trend to that of energy declining from 210, to 187 and finally 137 ml.l-1 of body pool. The amount drunk on visits to the watering point at daily, 2- and 3-day intervals was 281, 181 and 521 respectively.
There are few field data with which to validate this modelling of different pastoral watering regimes. As far as weight loss is concerned, unimproved Boran cows under drought conditions in the Kenya highlands lost up to 15% bodyweight during the course of a 36-week lactation when supplied with water and hay supplement (Lampkin and Lampkin, 1960a; 1960b). Predicted values for the different components and sources of body water turnover look realistic. An exception could be the requirement to tolerate 21% dehydration (i.e. 2 × WD), and still produce 1 kg of milk a day. The weakest link in the chain of calculation could be the water cost of forage intake (Iw).
It is concluded that the calculated values are near enough to the real values. They reveal the comparative effects of the different watering regimes, and their objectives. The change from daily to 2-day watering is an attempt to maintain production for man, whereas 3-day watering is a survival strategy for the cow. By then, the animal has practically exhausted its reserves of fat, having used about 12.5 kg. It has also used about 12.5 kg of its original 95 kg of muscle, liver and gut wall. Therefore whatever equilibrium it has achieved is extremely precarious. In this weakened state of ambulatory aestivation, an energy deficit of only a few MJ ME may cause the animal to collapse from exhaustion during the period of greatest stress in the management cycle. This period occurs on the third day without water when the cow has to walk the 22 km to water and back with scant prospect of finding forage. Thirst may keep the animal on its feet until it has drunk, but there may be little incentive to keep going after that.
The camel replaces the cow as the main milk producer at or below 200 mm mean annual rainfall (Western, in press). Its physiological adaptations have been referred to in many of the preceding sections. Briefly, they include slow absorption of water from the rumen, thereby reducing the amount excreted by the kidneys after drinking; good reabsorption of water from faeces and kidneys; reabsorption of nitrogen by the kidneys and its recycling in the body; and a diurnal range in body temperature which increases from 2°C in the hydrated animal to 6°C in the dehydrated state.
The camel has a tolerance of dehydration of at least 27% of its bodyweight (Schmidt-Nielsen, 1965). Its long neck enables it to reach the lower branches of most trees and, unlike the giraffe, it also has no difficulty feeding on dwarf shrubs at ground level. Camels also eat good quality standing hay (Aristida spp.), and salt bushes (Sueda spp.) to offset sweating when there is a salt defficiency in the diet (Gauthier-Pilters, 1974; Williamson and Payne, 1978; C.R. Field, 1978 and unpublished). The camel population in Africa appears to be increasing, possibly because desertification is extending into large areas south of the Sahara (UNEP, 1977) and opening up new habitats for the camel. There is also a growing demand for camel meat (C.R. Field, unpublished).
It follows that the staple food of pastoral people in arid regions of Africa is camel's milk. Among the Rendille in northern Kenya, for example, milk constitutes 60 to 90% of the diet, and 80 to 90% of this milk comes from camels (Schwartz, 1979). Young men herding camels away from the household may even have to rely on camel's milk to provide their water needs for up to 1 month at a time (G. Powys, unpublished).
Milk production is more important for these pastoralists than a high level of calf production. In fact, they compete with calves for milk, which is possibly why calf mortality rates of up to 40% are recorded (Bremaud, 1969; Spencer, 1973; Dahl and Hjort, 1976; Williamson and Payne, 1978; Wilson, 1978a).
The male camel is also important in the pastoral way of life as a water carrier for the household. The number of baggage camels dictates the amount of water that can be fetched and hence the distance a settlement can be from water; the further it is away, the more forage there will be available and hence the more productive the livestock will be, particularly the lactating females (Table 32). Despite the important role of male camels, short-term human needs take precedence over the longer term interests of the settlement to raise healthy stock. Male camel calves are first deprived of milk; then as juveniles they are bled to the point of exhaustion, providing up to 35 l of blood per annum, presumably mainly at the end of the dry season; and at 3 to 4 years they are castrated to make them more amenable. If they survive to become water carriers they are attached to the settlement and work long hours before being let out to browse in the immediate environment, which may have been stripped of vegetation (Schwartz, 1979). The same problem of water carrying does not occur in cattle-herding communities because the grazing must be better to support cattle, and the distance to water shorter. Donkeys can therefore be used as water carriers, and rarely have to supply an additional nutritional role.
Table 32. Relation between number of loading camels per household, maximum settling distance from a single water source and potential area of utilisation in northern Kenya.
|
No. of loading camels per household |
nil |
0.25 |
0.5 |
1 |
|
Frequency of water transport |
daily |
daily |
2 days |
4 days |
|
Maximum settling distance (km) |
2.5 |
5 |
12.5 |
27.5 |
|
Potential area of utilisation (km2) |
177 |
314 |
962 |
3318 |
Source: Schwartz (1979).
Even less information is available on the camel than on the zebu cow, so extra groundwork has to be done and assumptions made before the energy and water economy of the animal can be modelled.
As far as the energy budget is concerned, forage DMI has been found to vary between 1.4 and 2.5% of bodyweight (C.R. Field, unpublished); the gross energy and metabolisability of the camel's diet, which is mainly browse, has been calculated using the same coefficients as for grass. The following assumptions have been made about energy expenditure: cost of feeding and lactation are the same as for cattle, but camels are faster and more efficient walkers.
The components of TBWT can be built up in much the same way as for cattle (section 5.1.8), once the three coefficients, MYw, WKw and Iw have been estimated. The water cost per kg of milk (MYw) is taken as the same as for cattle, namely 0.87 l.kg-1. The likelihood of an extra water cost from walking in the sun (WKw), apart from that incurred foraging (Iw), has been disregarded in the management situations which will be illustrated. Therefore the only remaining, and most important, coefficient to be determined is the water cost per kg DMI (Iw) at specified environmental temperatures. No specific experiments have been designed to discover the value of Iw, but two pieces of fieldwork in Kenya can be used to arrive at estimates from which a mean figure can be taken.
Table 33. Data for estimate of water cost of forage intake in camels.
|
Item |
Unit |
n |
Mean |
S.E. |
|
C |
|
|
| |
|
Mean ambient temperature |
°C |
3 |
25.4 |
1.00 |
|
Moisture in diet |
fraction |
3 |
0.36 |
0.003 |
|
Water drunk (WD) |
l.d-1 |
8 |
12.9 |
0.86 |
|
Total body water turnover (TBWT) |
l.d-1 |
8 |
20.2 |
0.86 |
|
Water turnover rate constant (k) |
fraction |
8 |
0.006 |
0.0026 |
|
Bodyweight |
kg |
8 |
430 |
19 |
|
K |
|
|
| |
|
Period between drinks |
days |
|
17 |
|
|
Bodyweight |
kg |
|
500 (est.) |
|
|
Maximum amount drunk |
l |
|
100 (est.) |
|
|
Water drunk (WD) |
l.d-1 |
|
5.9 (est.) |
|
|
Milk yield(MY) |
kg.d-1 |
|
6 (est.) |
|
|
Moisture in diet |
fraction |
|
0.7 (est.) |
|
|
Forage intake(I) |
kg |
|
12.5(est.) |
|
Source: J.M. King et al (unpublished); C.R. Field (unpublished).
In the first trial, on Galana ranch, tritiated water turnover and related measurements were made on eight camels during June and August, the long dry season. The animals were male or dry non-pregnant females and so were treated as one group. The equation to be solved is (5.15), for which only WD has been measured (Table 33):
Iw = (WF + WD)/I l.kg-1DM.d-1
However water from food (WF) can be roughly estimated from equation (5.19):
WF = TBWT - (WD + WM + WRC) l.d-1
where
TBWT = 20.2 (Table 33)
WD = 12.9 (Table 33)
WM = 2.2 (Table 33 and equation 5.13)
WRC = 2:2 (Table 33 and equation 5.14)
Food intake (I) can be even more roughly estimated from equation (5.20) rewritten as:
I = WF (1- forage moisture)/forage moisture
I = 2.9 (1 - 0.36)/0.36
I = 5.16 kg DM.d-1
Equation (5.15) can now be solved:
Iw = (2.9 + 12.9)/5.16
Iw = 3.06 l.kg-1DM.d-1
The value for Iw could be an overestimate because it is based on a forage DMI of only 1.2%, whereas at the minimum DMI measured (C.R. Field, unpublished) the value for Iw would be 2.6.
In the second trial near Mount Kulal in northern Kenya, lactating camels walking 2 km.d-1 in dense green vegetation went without drinking for 14 to 20 days (C.R. Field, unpublished). Given this information a number of assumptions can be made based on measurements of the assumed values in that or similar environments (Table 33). The equation to be solved for Iw is, in this case, (5.21)
Iw = (WD + WF) - (WK × WKw) - (MY × MYw)/I.
Figures for some of these variables can be substituted directly from Table 33:
IW = (5.9 + WF) - 0 - (6 × 0.87)/I
Values for intake of very green vegetation by lactating camels are likely to be at the top of the recorded range, namely 2.5%. Given this information, WF can be solved (equation 5.20):
WF = (12.5 × 0.7)/(1 - 0.7)
WF = 29.2 l.d-1
Reverting to equation (5.21):
Iw = (5.9 + 29.2) - (6 × 0.87)/12.5
Iw = 2.4 l.kg-1DM.d-1
The mean figure for Iw from the two studies is 2.7 l.kg-1DM.d-1, and this will be used in the following illustrations.
The purpose of the examples is to demonstrate the effect of the application and removal of constraints on energy and water use in the lactating camel. In the first situation the camel is being kept in a poor settlement, 5 km from water, to provide milk for the women, children and old people. In the second, the same animal has been released to complete the second half of its lactation in the mobile herd, which is managed by boys and young men (Dahl and Hjort, 1976).
The basic data required to model the energy budget and water turnover of the animal are given in Table 34.
Table 34. Basis for calculating the daily energy budget and water turnover of a camel in two different situations.
|
Item |
Unit |
Settlement 5 km from water |
Mobile herd |
|
Energy budget |
|||
|
Expected liveweight (WE) |
kg |
500 |
500 |
|
Forage intake |
%WE |
1.9 |
2.4 |
|
kgDM |
9.6 |
11.8 |
|
|
Forage digestibility |
fraction |
0.45 |
0.55 |
|
Actual liveweight |
kg |
500 |
460 |
|
Distance walked |
km |
16 |
16 |
|
Walking cost |
kJ.kg-1.km-1 |
1.0 |
1.0 |
|
Milk yield |
kg |
5.0 |
4.0 |
|
Milk energy |
MJ.kg-1 |
3.5 |
3.5 |
|
Water turnover |
|||
|
Forage moisture |
l.kg-1 |
0.35 |
0.64 |
Source: Nutritional analysis by University of Hohenheim of dietary grab samples collected by C.R. Field, who also measured DMI, distances walked and daily activity; Schmidt-Nielsen et al (1967); Dahl and Hjort (1976); Knoess (1977); Williamson and Payne (1978); King (1979).
Table 35. Estimated daily energy budget and water intake of a camel in two different situations.
|
Item |
Energy budget (MJ ME) |
|
|
Settlement 5 km from water |
Mobile herd |
|
|
Forage intake |
63.0 |
94.6 |
|
Maintenance at rest |
33.5 |
30.4 |
|
Extra costs: feeding |
2.5 |
3.8 |
|
walking |
8.0 |
7.4 |
|
lactation |
31.8 |
25.5 |
|
Total expenditure |
75.8 |
67.1 |
|
Energy balance |
-12.8 |
+27.5 |
|
|
Water intake (l) |
|
|
Forage moisture |
5.2 |
21.0 |
|
Drink |
25.1 |
14.3 |
|
Metabolic |
2.2 |
1.9 |
|
Respiratory and cutaneous |
3.9 |
4.5 |
|
Total input |
36.4 |
41.7 |
Source: Table 34, section 5.1.
The results of the calculations are presented in Table 35. Looking down the first column of figures, it can be seen that the camel attached to the settlement is losing weight at 2.9 kg per week. Any energy deficit that the animal might be expected to have during the first half of its lactation is accentuated by the lack of forage accessible to the camel from the settlement. At this proximity to permanent water, the vegetation will almost certainly be overused by livestock. It is, of course, man and not the animal who has to stay near water; the camel only needs to drink every 4 days. After 3 months at the settlement the animal will have lost about 40 kg, i.e. 8% of its bodyweight, and might then be moved to the mobile herd.
The mobile herd provides most of the food and water needs of the herdsmen, and so it is not tied to the watering place by man's drinking water requirements. The herd therefore can exploit the more inaccessible parts of the country where the vegetation is of better quality and greener, unless the herding range is restricted by intertribal fighting (Field, 1978). The camel, introduced to this herd from the settlement, is able to revert to a positive energy balance. The rate of weight gain is calculated from equations (5.08) and (5.09) as 0.8 kg per week, so that within approximately 7 weeks it will have regained its original weight. At the same time its water turnover rate has increased, but its drinking requirement has decreased due to the moisture content of the forage. It would need to drink once in 7 days.
The above calculations of the rate of water use by camels can be compared with data from C.R. Field (unpublished) who found that camels drank 100 to 1351 of water every 6 to 7 days when attached to a settlement.
Sheep and goats are the 'petty cash' of livestock capital, and are used by man to fulfill a variety of different social and religious obligations. They are also an insurance against a dry year because smallstock, and in particular goats, may outsurvive cattle. Even if it is not so dry that cattle survival is in doubt, it is likely that cows' milk will have dried up and people will have resorted to bleeding male animals and using smallstock (Dahl and Hjort, 1976). Sheep and goats will be slaughtered for food, or sold to buy maizemeal. Goats, in particular, will be expected to give milk under the driest conditions (A.C. Field, 1978).
Smallstock exploit a broader spectrum of natural bushed grassland, and a wider range of rainfall than cattle (Knight, 1965; Topps, 1967). Sheep and goats are usually herded by women and children nearer to the homestead than zebu cattle and camels, but they can cover considerable distances when required to do so (Dahl and Hjort, 1976). Bedouin goats often walk 17 km.d-1 for grazing and return to the encampment every evening to suckle their kids. The encampment may be 25 km from the watering point (A. Shkolnik, unpublished). Australian sheep covered 15 km.d-1 at 2 to 2.5 km.h-1 when food and water were separated (Squires and Wilson, 1971). As the distance of separation increased, food intake decreased almost linearly; at first water intake remained the same, but then it declined; the distance walked per day increased to a peak and then dropped to the initial level, and frequency of watering decreased (Squires, 1978).
Although the two species are herded together, that is where the similarity ends. They are kept for different purposes, and their products are not entirely interchangeable: for instance, sheep are kept for their fat and goats for their lean meat. They have different susceptibilities to different diseases (BVA, 1976), so by keeping both the pastoralist is spreading the risk. In arid areas, both species will take more than one third of their annual diet as browse and browse litter, but the composition of the diet differs.
In the lowlands of northern Kenya it was noted that sheep seldom raise their heads above 20 (cm from the ground when feeding, and goats normally do not lower their heads below 20 cm (IPAL, 1983). Goats can extend their feeding range by standing on their hindlegs (A.C. Field, 1978), or even climbing trees. Consequently they can ingest a higher proportion of trees and shrubs than sheep (Table 36). In fact the presence of goats is frequently associated with overgrazed, degraded areas. It has been pointed out that goats may not have been the cause of the original overgrazing (Devendra and Burns, 1970; Joubert, 1973). They are merely delivering the coup de grace: for example, Turkana people with goats can survive after Rendille and Gabra have grazed out an area with their camels and cattle (C.R. Field, unpublished).
Water balance is unaltered during the first 2 days of water deprivation in sheep, with water drawn from the digestive tract; only after that does physiological dehydration develop (Hecker et al, 1964). It reaches a critical level only when desert sheep have lost 30% of their bodyweight (More and Sahni, 1978), and at an even later stage in desert milking goats (Shkolnik et al, 1972). When deprived of water, sheep and goats reduce water loss by the excretion of sodium in concentrated urine (Macfarlane et al, 1961; Taneja, 1965; Schoen, 1968). Faecal water loss is also limited, particularly by breeds originating from arid areas (Slagsvold, 1970).
Differences in food selection and water requirements between breeds of the same species are nearly as large as differences between the two species in different parts of Africa: for example, compare Gihad (1976) and King (1979). Nevertheless the practice of grouping sheep and goats together under the heading of smallstock should be discontinued as more field data become available.
From the foregoing remarks it is apparent that it will be difficult to generalise from one chosen example. And data on water and energy budgets are so sparse that differences between species and breeds cannot be illustrated with confidence.
Table 36. Difference in the diet of sheep and goats in the dry season in northern Kenya.
|
|
Per cent of diet |
||
|
Plant species |
Type |
Sheep |
Goats |
|
Cordia sinensis |
Large shrub |
0 |
3 |
|
Acacia nubica |
Large shrub |
0 |
13 |
|
lndigofera spinosa |
Dwarf shrub |
21 |
39 |
|
Indigofera cliffordiana |
Dwarf shrub |
1 |
1 |
|
Hermannia kirkii |
Herb |
1 |
9 |
|
Leptothrium senegalense |
Grass |
14 |
7 |
|
Kohantia caespitosa |
Herb |
6 |
1 |
|
Total |
|
43 |
73 |
Source: A.C. Field (1978); C.R. Field (unpublished).
One animal which has been and is being studied, is the Small East African goat, which is common to all but the coldest or driest parts of the region.
The forage intake of the goat has been taken as 2.5% of bodyweight, based on a range of 2.1 to 3.2% (Devendra and Burns, 1970; A.C. Field, 1978). The gross energy and metabolisability of the diet has been calculated using the same coefficients as for grass; the cost of feeding has been made the same as for cattle; walking cost was taken as higher and cost of lactation as lower than for large stock, but based on a higher nutritional value (Dahl and Hjort, 1976) than that shown in Table 9.
The water cost of each kg of milk has been taken as the same as for cattle, and the extra water used in walking as negligible. The water required per kg DMI (Iw) at a given environmental temperature has been calculated from data on Small East African goats at Galana ranch, which were included in the field trials with the camels (Table 37).
Table 37. Data for estimate of water transactions in Small East African goats.
|
Item |
Unit |
n |
Mean |
S.E. |
|
Mean ambient temperature |
ºC |
4 |
25.4 |
0.66 |
|
Moisture in diet |
fraction |
4 |
0.53 |
0.118 |
|
Water drunk (WD) |
l.d-1 |
12 |
2.20 |
0.295 |
|
Total body water turnover (TswT) |
l.d-1 |
12 |
3.86 |
0.187 |
|
Water turnover rate constant (k) |
fraction |
12 |
0.155 |
0.0075 |
|
Bodyweight |
kg |
12 |
36.6 |
1.67 |
Source: J.M. King et al (unpublished).
Water from food has been derived from equation (5.19):
WF = TBWT - (WD + WM +WRC) l.d-1
where
TBWT = 3.86 (Table 37)
WD = 2.20 (Table 37)
WM = 0.22 (Table 37 and equation 5.13)
WRC = 0.42 (Table 37 and equation 5.14)
Hence WF was 1.02 l.d-1. The mean value for forage intake (I) during the period from June to August was taken as one third of the way up the range of observed values, namely 0.90 kg.d-1. The forage moisture content was then calculated as 53% from equation (5.20).
The water intake in relation to forage intake is estimated from equation (5.15) as follows:
Iw = (WF + WD)/I
Iw = (1.02 + 2.20)/0.90
Iw = 3.61.kg-1DM.d-1
This value compares reasonably well with the value for growing sheep under hot conditions, namely 3 l.kg-1DM.d-1 (Table 19), because the goats on Galana were not particularly arid-adapted, and the temperature was higher.
The application of the values derived above to the energy and water turnover of the goat is shown in Table 38. It can be seen that the animal' would have an energy deficit of 1.64 MJ ME.d-1, resulting in a weight loss of 0.38 kg per week. Such a rate of loss could be tolerated for about 3 months, assuming that forage intake was also decreasing. It is more likely that the animal would dry off before then and eliminate the reason for the deficit, which is the energy cost of lactation. Frequency of watering would need to be at least every third day to avoid depression of appetite and lactation (section 4.3.1).
The Small East African goat is not as desert-adapted as the Somali sheep or Galla goat, which only need to drink once a week when cattle are being watered every second day (Mares, 1954). Furthermore the goat has a higher TBWT than the zebu cow (Table 13). Nevertheless it possesses, in some degree, the attributes which allow smallstock to outsurvive cattle in times of drought. It requires less water per kg DMI than zebu cattle in the same environment. Its food intake is higher because of its superior ability to select nutritious food, except during the rains when there is adequate forage anyway. As a browser it obtains more water from forage than do grazers. Nevertheless in a severe drought, when the leaves turn brown and fall off even the deep-rooted shrubs, smallstock may lose their edge over cattle when faced with the same indigestible diet. They may be able to find natural supplements such as browse litter. For example, acacia flowers rich in protein and soluble carbohydrates are shed at the end of the dry season (Schwartz and Said, 1980).
Table 38. Daily energy budget and water intake of a Small East African goat.
|
Item |
Value |
Unit |
Coefficient |
Equation number |
Energy (MJ ME) | |
|
Forage intake |
0.90 |
kg DM |
d = 0.50 |
(5.02) |
6.40 | |
|
Maintenance at rest |
35 |
|
0.251 W0.75 |
(5.04) |
5.38 | |
|
Extra costs: |
|
|
|
|
| |
|
|
feeding |
6.40 |
MJ ME |
|
- |
0.26 |
|
|
walking |
8 |
km |
1.9 kJ.kg-1.km-1 |
- |
0.53 |
|
|
lactation |
0.35 |
kg |
3.0 MJ.kg-1 |
(5.05) |
1.87 |
|
Total expenditure |
|
|
|
|
8.04 | |
|
Energy balance |
|
|
|
|
-1.64 | |
|
|
|
|
|
|
Water intake (I) | |
|
Forage moisture |
0.50 |
fraction |
- |
(5.20) |
0.88 | |
|
Drink |
- |
- |
Iw = 3.61.kg-1 DM |
(5.21) |
2.40 | |
|
Metabolic |
8.04 |
MJ ME |
- |
(5.12) |
0.23 | |
|
Respiratory and cutaneous |
- |
- |
- |
(5.22) |
0.42 | |
|
Total input |
|
|
|
|
3.93 | |
Source: Clapperton (1961); Young (1966); Graham (1964); Corbett et al (1969); Devendra and Burns (1970); McDonald et al (1976); A.C. Field (1978); Webster(1978); King(1979); nutritional analysts by University of Hohenheim of dietary grab samples collected by C.R. Field.
The donkey has been described as the ubiquitous beast of burden in southern Sudan (Wilson, 1978b); a role that it also fulfills in much of sub-Saharan Africa. In subsistence societies which value multipurpose animals, the retention of a specialist animal is surprising. Equines are no longer milked in Africa as they are in Asia (Jarrige, 1980), and donkeys are eaten only as a last resort. The animal owes its survival to the following attributes: it can carry a heavier load in relation to its bodyweight than other baggage and riding animals (Table 39); it is more efficient at walking than man, or even than the mule (Yousef and Dill, 1969); it appears to thrive on neglect; and it can withstand droughts better than most species of livestock, as was illustrated in Niger in the early 1970s (Eddy, 1979). The characteristics which enable it to survive are summarised below.
Table 39. Daily performance figures of baggage and riding animals.
|
Animal |
Bodyweight (kg) |
Job |
Load to bodyweight ratio |
Load (kg) |
Natural speed (km.h-1) |
Distance travelled (km) |
|
Donkey |
125c |
Pack |
0.60b |
75 |
3.2g |
16e |
|
|
160c |
Ride |
0.50 |
80e |
4.0g |
20e |
|
Horse |
350b |
Ride |
0.23 |
80e |
8.0e |
30e |
|
Camel |
450a |
Pack |
0.51 |
230d |
4.0d |
24d |
|
|
450a |
Ride |
0.27 |
120d |
10.0d |
48d |
|
Ox |
350b |
Pack/Ride |
0.31f |
110e |
5.5b |
20e |
Source: aWilson (1978a); bCEEMAT (1972); cWilson (1978b); dWilliamson and Payee (1978);eestimate; fIshizaki et al (1961); gE.M. Wathuta (unpublished).
African equines exploit the rangelands in a very different way from the ruminant. Their food intake is not depressed by a low protein, high fibre diet (Robinson and Slade, 1974), and so the donkey remains productive with little loss of condition. The concomitant demands for water for cooling and for intermediary metabolism are high. The working donkey uses water at much the same rate as man (Dill, 1938), but the water that it carries back to the boma is for the household and not for itself. Fortunately the animal is more adept than man at maintaining its plasma volume at the expense of other extracellular and intracellular fluids. Its appetite is not depressed until dehydration is severe, by which time the donkey is back at the watering point where it rapidly re-hydrates without risk of intoxication (Schmidt-Nielsen, 1965; Maloiy, 1970; Maloiy and Boarer, 1971). When the animal is not working, minimum water expenditure could be as low as 2.5% (Dill, 1938), which would allow the animal to go without drinking for at least 10 days, because it can tolerate 30% dehydration. But donkeys in northern Kenya appear to water more frequently than smallstock (C.R. Field, unpublished).
Donkeys are used to carry grain, forage, firewood, people, household possessions and water, as well as to pull carts. In Maasailand, the animal usually weighs about 120 kg and carries a load of 35 to 45 kg. In the dry season the location of a boma may, in part, be determined by the distance a donkey can walk to a source of drinking water and back to the household. The journey should be completed during daylight, i.e. within 13 h at speed of about 3 km.h-1 (Table 39). Thus a settlement can be up to 20 km from a source of drinking water, end a donkey will be expected to make the trip there and back every second or third day during the dry season (D. Western, unpublished). On the days it is not working the donkey may follow the cattle out to graze, but go less far, and be brought back to the vicinity of the boma in the evening. It is not enclosed at night unless there is a high risk of predation.
The example chosen is less extreme than the limits of endurance credited to the donkey would permit and is more in line with the 2-day watering regime of zebu cattle, where the boma was located 1 km from water (section 5.2.1). When the animal is being used to collect water on alternate days, its daily TBWT and energy budget will be approximately as shown in Table 41, based on the coefficients and equations listed in Table 40.
The first point to note is the high estimate of forage intake (2.7% of bodyweight) despite the fibrous diet. The donkey being a simple-stomached herbivore, achieves microbial digestion of plants in its large intestine, which has several functional similarities to the rumino-reticulum, including the synthesis and digestion of microbial protein (Stevens et al, 1980). The efficiency of cell-wall digestion of forages is 30% less in Equidae than in ruminants, while the apparent digestibility of the cell contents is similar. The digestibility (d) of forage can be predicted from either of the following equations, or preferably the mean of the two:
d = 0.31 + 1.45 CP (5.23)
where CP = crude protein fraction, or
d = 1.45 - 2.82 CF (5.24)
where CF = crude fibre fraction.
The reduced breakdown of cell-wall carbohydrates in the large intestine of Equidae is primarily due to a shorter retention time than in ruminants. The latter must retain fibrous vegetation in the rumen until it is small enough to pass through the reticulo-omasal orifice and between the leaves of the omasum. This process may be so delayed that food intake falls below the level required for maintenance regardless of the quantity of herbage on offer. No such depression of intake occurs in the equine which compensates for its less efficient digestion with a 30% longer feeding time than the ruminant. The conclusion is that caecal digestion is in fact a superior adaptation for dealing with high fibre content herbage, provided that intake is not limited by the actual quantity of herbage available. Amongst the Equidae, the donkey appears to have a higher capacity for crude fibre intake than the horse (Robinson and Slade, 1974; Jarrige, 1980). For example, the donkey is one of the few species that can use the short spiky leaves of Sporobolus spicatus, a common grass of alkaline flats in Africa (Pratt and Gwynne, 1977).
The metabolisability of the food digested is slightly higher (0.90) than for ruminants, so that the energy derived (E) from a given intake (l) is predicted with a different equation than equation (5.02). For equines:
E = I × 18 × d × 0.90 MJ ME (5.25)
The efficiency (km) with which ME is used for maintenance has been derived from an adaptation of equation (5.04):
km = 0.55 + (0.3 × 0.90 × d) (5.26)
Table 40. Estimates of daily energy budget and water turnover of a donkey carrying water on alternate days.
|
Item |
Coefficient or equation |
Value |
Unit |
|
Energy budget |
|||
|
Bodyweight (Wt) |
- |
120 |
kg |
|
Forage intake (I) |
0.027 Wt |
3.2 |
kg DM |
|
Crude protein (CP) |
- |
0.06 |
fraction |
|
Crude fibre (CF) |
- |
0.42 |
fraction |
|
Forage digestibility (d) |
(5.23/4) |
0.33 |
fraction |
|
ME intake |
(5.25) |
17.1 |
MJ ME |
|
Efficiency of ME use (km) |
(5.26) |
0.639 |
fraction |
|
Maintenance cost at rest |
(5.26/7) |
12.0 |
MJ ME |
|
Feeding cost |
53 kJ.MJ ME |
0.9 |
MJ ME |
|
Walking unloaded (including grazing) |
1.0 kJ.kg-1 bodyweight.km-1 |
11.5 |
km |
|
Walking loaded |
1.4 kJ.kg-1 bodyweight.km-1 |
5.5 |
km |
|
|
Water turnover |
||
|
Forage moisture |
(5.20) |
0.1 |
fraction |
|
Water cost relative to food intake (Iw) |
3.6 l.kg-1DM |
11.5 |
l |
|
Walking speed |
- |
3.2 |
km.h-1 |
|
Distance between water and boma |
- |
11 |
km |
|
Time walking loaded and hot |
- |
1:8 |
h |
|
Water cost, walking loaded and hot (WKLw) |
0.0133 Wt |
1.6 |
l.h-1 |
|
Metabolic water |
(5.12) |
0.5 |
l.d-1 |
|
Respiratory and cutaneous water |
(5.22) |
1.6 |
l.d-1 |
Source: Dill (1938); ARC (1965; 1980); Fonnesbeck (1968); Vander Noot and Gilbreath (1970); Wooden et al (1970); Hintz et al (1971); Vander Noot and Trout (1971); CEEMAT (1972); Robinson and Slade (1974); Schmidt-Nielsen (1975); Pratt and Gwynne (1977); E.M. Wathuta (unpublished).
Thus the amount of ME used for maintenance by the donkey is given by the equation:
Em = a.Wt0.75/km (5.27)
where a = 0.211 (Table 24).
The energy costs above maintenance are feeding and walking with and without a load. Feeding costs per MJ ME have been increased by one third to account for the extra feeding time. Walking costs have been based on the assumption that the donkey walks only 3 km while grazing on the day it is carrying water but 9 km on the day it is not. To this total must be added the 11 km journey to water carrying empty containers. The load on the 3.5 h return journey has been estimated at 45 kg, i.e. less than 40% of the animal's bodyweight, which is not hard work for a donkey. Larger loads and longer working hours will increase energy requirements by two to four times that of maintenance (CEEMAT, 1972; BDPA, 1974).
Water intake from forage is low, because the grass is dry. Drink therefore accounts for most of the animal's water intake. It is calculated using a variation of equation (5.21), namely:
WD = (I × Iw) + (WKU × WKUw) + (WKL × WKLw) - WF (5.28)
where the water required per kg DMI (Iw) has been taken at the same value as for the light horse at a moderate ambient temperature (15°C) (Table 40).
The water cost of walking unloaded (WKUW) has been taken as negligible, for two reasons. Because the donkey is an efficient walker it generates relatively little heat and there is no marked increase in sweating in response to exercise, as there is in man. Speed of walking is dictated by the animal's ability to maintain thermoregulation, and it will not be driven any faster. Thus the inherent stubborness of the animal enables it to avoid working at a rate exceeding its thermoregulatory capacity (Yousef and Dill, 1969; Bullard et al, 1970). Secondly, the animal will have covered the distance to water before the day heats up.
There is a water cost of walking back loaded (WKIw) during the heat of the day of about 1.61.h-1 (Table 40). If the return journey is not delayed until the afternoon this cost will be borne by the animal for 3.5 h, which must be divided over 2 days. Equation (5.28) can be solved:
WD = 11.5 + 0 + 2.8 - 0.4
WD = 13.9 l.d-1
Estimates of metabolic and respiratory and cutaneous water input have been made using equation (5.12) and (5.22) respectively.
Table 41. Estimated mean daily energy budget and water intake of a donkey carrying water on alternate days.
|
Activity |
Energy budget |
|
MJ ME |
|
|
Forage intake |
17.1 |
|
Maintenance at rest |
12.0 |
|
Extra costs: feeding |
0.9 |
|
walking unloaded |
1.4 |
|
walking loaded |
0.9 |
|
Total expenditure |
15.2 |
|
Energy balance |
+ 1.9 |
|
|
Water intake (l) |
|
Forage moisture |
0.4 |
|
Drink |
12.4 |
|
Metabolic |
0.5 |
|
Respiratory and cutaneous |
1.6 |
|
Total input |
14.9 |
Source: Table 40.
The final picture of energy and water turnover in the donkey (Table 41) is very different from that in cattle belonging to the same household, also on a 2-day watering regime (Table 31). The donkey has an energy surplus whereas the cow has a deficit. The reason appears to be that the donkey has a high ME intake, which in the ruminant would be associated with high energy demands, for example in lactation, whereas in fact the donkey's maintenance requirements are very low.
However, water expenditure (ml.l-1) is similar in both animals. There are two reasons for this: firstly, although the water cost per kg DMI (Iw) is lower in the donkey, food intake must be higher for a given ME intake; secondly, the evaporative water needs of the donkey walking in the heat of the day are nearly three times those of the cow walking unloaded and more than offset the cow's extra water need for lactation.
The rate of water loss in the donkey is particularly marked at the outset of the 2 days between drinking, because of the sweat lost carrying the water back to the boma. This loss must not reach a level which will depress the animal's appetite during the next grazing day, even though it may be able to graze during the first night. On the morning after work, the degree of dehydration will be approaching 13% and by the afternoon 15%. At this stage the person who has walked with the donkey, and sweated nearly as much, would be in a coma if he or she had not drunk. However, donkeys are relatively unaffected by a water loss of 12 to 15% of their bodyweight; appetite is not depressed until the level of dehydration reaches in excess of 15%, or as high as 23% in Sinai donkeys (A. Shkolnik, unpublished). By the time the animal returns to drink on the second day it will have lost 21% of its body water, but will rapidly and completely rehydrate (section 3.3.1).
The foregoing examples and equations have been based largely on data and situations in Kenya. Nevertheless, the approach is readily transferable to other parts of Africa. For example, given baseline data of the sort provided by Swift (1979) on the Tuareg in Mali (Table 42), water and energy turnover of four species can be simulated.
The value of the model is that the behaviour of livestock throughout the year can be illustrated and related to a variety of options for water development, whereas practical guidelines for development were formerly based on a single conservative estimate of daily drinking water requirements (Table 10).
Table 42. Distribution of nomadic flocks and herds in relation to water during dry seasons in arid Mali.
|
Species |
Water frequency (d) |
Distribution of flocks and herds (%) |
|||
|
Distance between pasture and wells (km) |
|||||
|
10-19 |
20-29 |
30-39 |
40+ |
||
|
Camels |
5 - 6 |
25 |
23 |
39 |
13 |
|
Cattle |
2 - 3 |
19 |
66 |
15 |
0 |
|
Sheep |
3 |
30 |
54 |
16 |
0 |
|
Goats |
- 2 |
54 |
46 |
0 |
0 |
Source: Swift (1979).
![]()
![]()
![]()
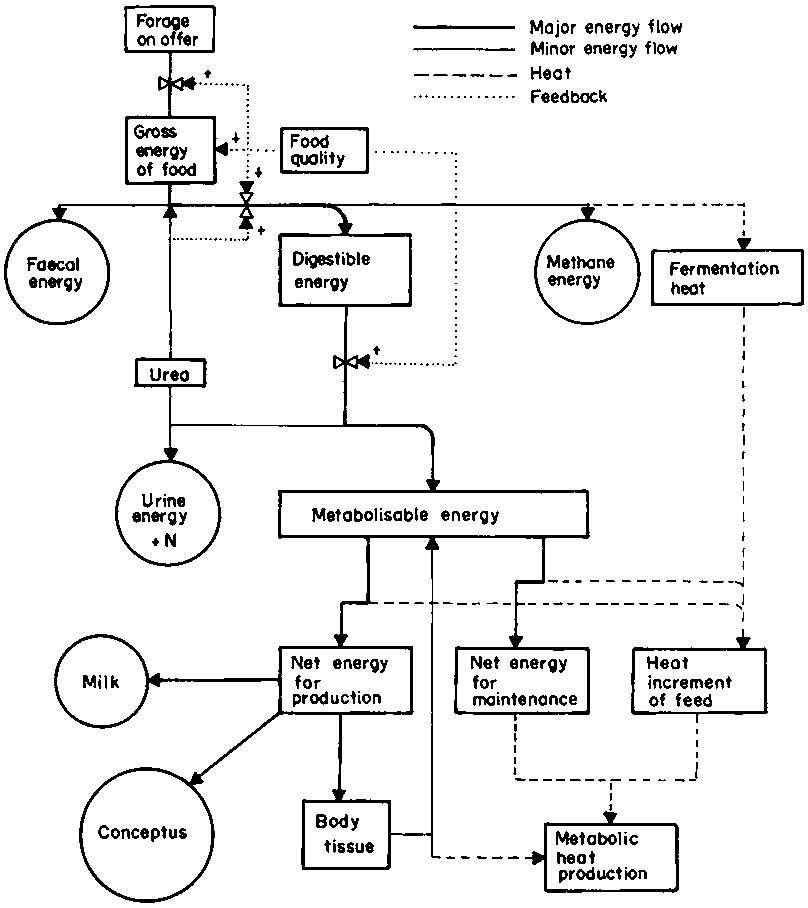{kind=link}
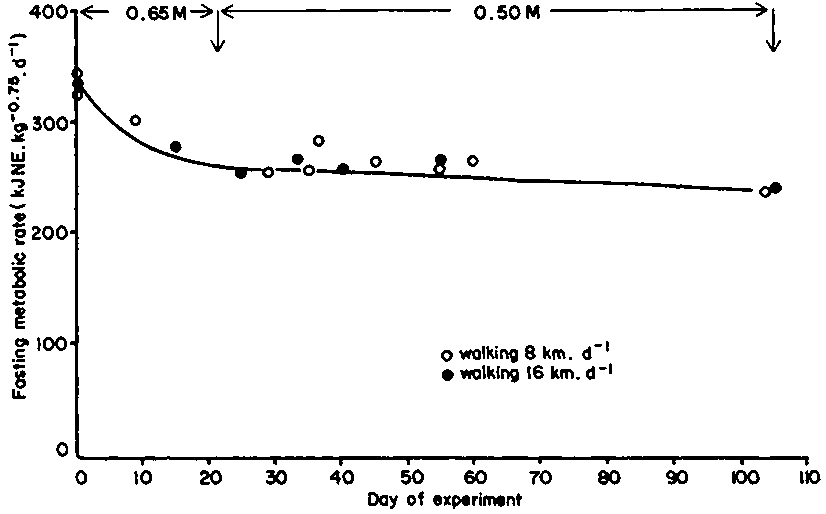{kind=link}
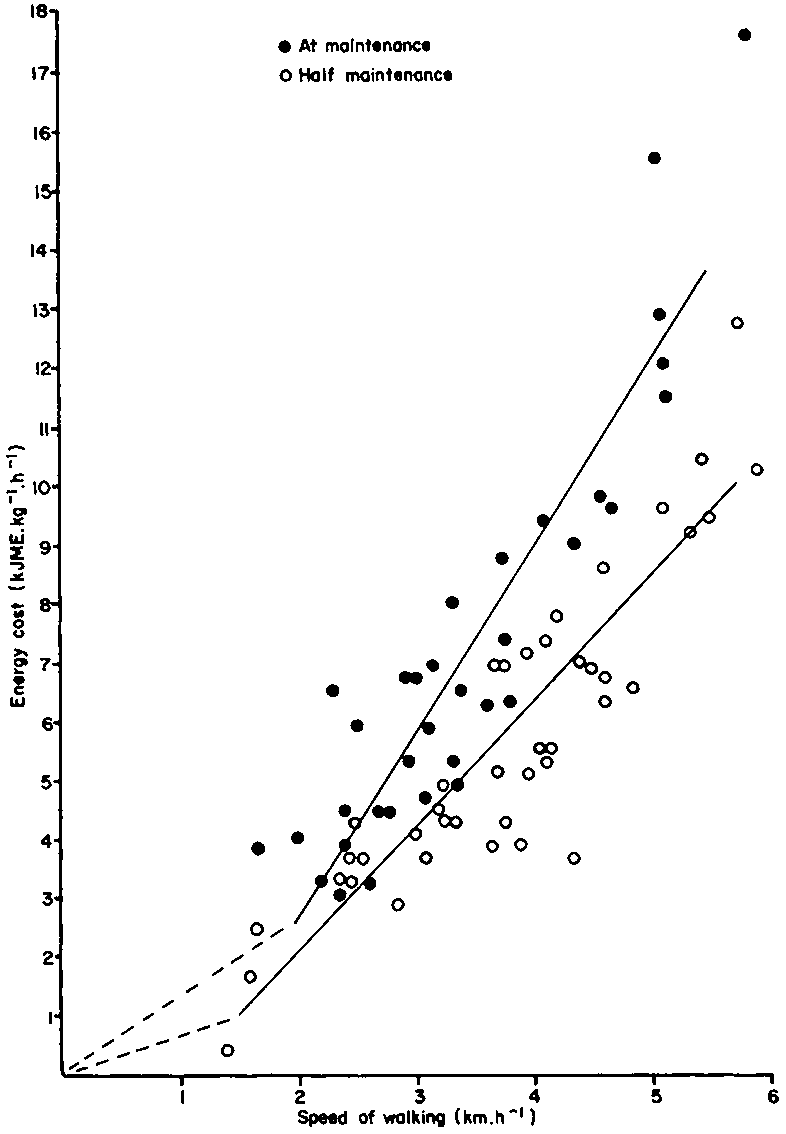{kind=link}