![]()
![]()
![]()
Extent of the problem
Complexity of the problem
Control of the problem
The current world human population is estimated to be 4.2 billion and is expanding by about 2% or 70 to 80 million people each year (Gavora, 1982). To deal with this increase, an additional 30 million tonnes of staple foods will be needed annually. A major way in which livestock production can be increased, and thus help meet this need, is the reduction of livestock losses due to disease. It is estimated that a 2% reduction in livestock losses due to disease would provide food for an additional 80 million people (Gavora, 1982).
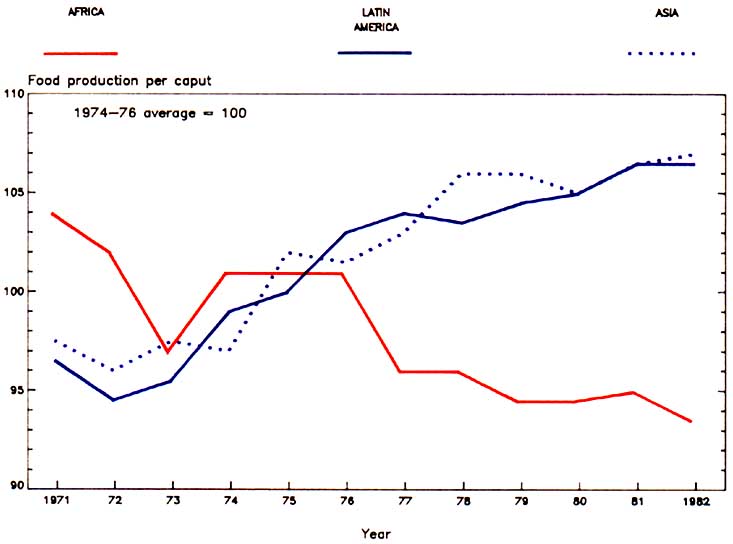
The situation is most serious in developing countries where 75% of the world's population lives. It is estimated that 65 to 70% of the world's livestock resources exist in these regions, yet they account for only 30% of the world's meat output (FAO, 1975). Over the last 5 years, Africa's population has increased at twice the rate of food production and by the year 2000 will have risen from the current estimated 470 million to 877 million (Anon, 1984). In fact food production per capita in the continent is falling compared with an overall increase in Asia and Latin America (Figure 1).
Figure 1. The decline in food production per caput in Africa.
The production per unit area of animal protein is lower in Africa than in any other continent. For Africa, the estimated production of animal protein from livestock farming per 1000 hectares is 542 kg, for Latin America 4113 kg and for Europe 38083 kg (FAO, 1975). However, Africa has vast agricultural resources which are at present underdeveloped and untapped.
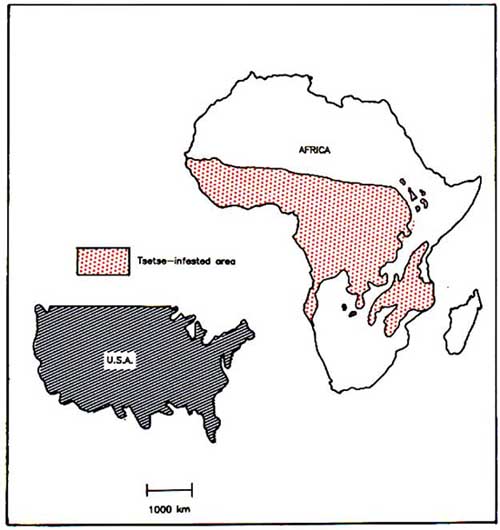
The causes of this serious deteriorating situation are complex, but probably the single most significant factor is the animal African trypanosomiasis group of diseases transmitted by tsetse flies. Currently, vast humid and subhumid areas of Africa are held captive by tsetse flies and the trypanosomes which they transmit. Tsetse flies infest 10 million km2 of Africa (Figure 2), representing 37% of the continent or about half the habitable land, and affect 37 countries (FAO/WHO/OIE, 1982). It is considered that 7 million km2 of this area would otherwise be suitable for livestock and mixed agriculture. It is conservatively estimated that this area could support another 140 million cattle plus at least equivalent numbers of sheep and goats. Much of the best watered and most fertile land is infested with tsetse, while large areas of good grazing, most notably in the subhumid zone, could be immediately used by pastoralists if trypanosomiasis could be controlled (MacLennan, 1980).
Figure 2. Tsetse flies infest an area of Africa larger than the USA.
Currently, about 30% of the 147 million cattle in countries affected by tsetse are exposed to infection (FAO/WHO/OIE, 1982). The situation for sheep, goats, pigs, horses, donkeys and camels is probably similar but is less well documented (Table 1). The presence of tsetse flies not only excludes domestic livestock from a considerable area of Africa but also causes severe losses in livestock production due to poor growth, weight loss, low milk yield, reduced capacity for work, infertility and abortion. The annual loss due to tsetse in meat production alone is estimated at US$ 5 billion (FAO/WHO/OIE, 1963). Further losses occur in milk production and in mixed agriculture where draught power and manure play a vital role. In Africa, only 20% of tractive power is mechanised (McDowell, 1977). It has been estimated that a draught ox can increase the agricultural output of a family unit six-fold (McDowell, 1977). Furthermore, the manure provided by livestock is essential for the production of both food crops and cash crops and is a source of energy in the form of biogas (McDowell, 1977). If all these factors are taken into consideration, it is estimated that the development of livestock and agriculture in tsetse-infested Africa could generate a further US$ 50 billion annually.
Table 1. Livestock populations in the 37 countries of Africa infested with tsetse fly.
|
No. of animals in tsetse-infested Africa (millions) | ||||||
|
Cattle |
Sheep |
Goats |
Pigs |
Horses |
Mules |
Camels |
|
147 |
104 |
125 |
8 |
3 |
9 |
11 |
Source: FAO/WHO/OIE (1982).
Countries in West and central Africa especially are severely affected by trypanosomiasis. Twenty six per cent of Africa's human population live in the 18 countries from Senegal across to Zaire, but this vast area maintains only 9% of Africa's cattle, sheep and goats (ILCA, 1979). As a result, the average livestock biomass per inhabitant in West and central Africa is only 26 kg in contrast to 136 kg for the remainder of Africa south of the Sahara and 79 kg for the continent as a whole. FAO (1974) estimates the average potential carrying capacity in this region to be 20 cattle/km2 as compared with the current 3.4 cattle/km2. Equivalent increases would also be possible for sheep and goats, i.e. approximately five times as many animals could be maintained. The situation in southern and East Africa is no better, with up to 70% of the land in some countries being infested by tsetse (e.g. Tanzania). It should be emphasised that approximately 50% of the domestic livestock in the 37 countries infested by tsetse are confined to six countries in East Africa (Table 2) and that this is only possible because a large proportion of these animals exist in highland areas, above the limits of tsetse infestation.
Available data indicate that the overall situation with regard to African trypanosomiasis in domestic livestock is deteriorating. MacLennan (1980) states that since the 1950s territorial expansions of tsetse infestation in savanna areas have been prodigious and are continuing, despite regression in some areas as a result of insecticide control programmes and increased cultivation and hunting by expanding human communities. Some of the more recent advances include an area of 26000 km2 in Nigeria, while in Central Cameroon an advance of Glossina morsitans has occupied about 21000 km2 and, unless halted, will proceed to occupy a further 9000 km2 of land which at present is extensively used for agriculture. In Zambia, tsetse has extended over 11700 km2 in the southwest. The full extent of tsetse spread is not known but other significant advances have been observed in Botswana, Zimbabwe, Malawi, Uganda, Tanzania, Sudan, Ethiopia, eastern Senegal and western Mali. The greatest and most active advance of modern times is probably proceeding in southern Angola (FAO, 1979; MacLennan, 1980).
This situation is resulting in increasing pressure on tsetse-free pastures and is bound to lead to pasture degradation, a fall in output and increased production costs. Thus, the argument that the tsetse conserves Africa is not valid, and it is becoming imperative to make fuller use of tsetse-infested areas to alleviate the pressure on tsetse-free regions.
Table 2. Populations of domestic ruminants in the East Africa region.
|
Country |
No. of domestic ruminants (millions) |
||
|
Cattle |
Sheep |
Goats |
|
|
Kenya |
11.5 |
4.7 |
4.6 |
|
Tanzania |
12.7 |
3.9 |
5.8 |
|
Uganda |
5.0 |
1.8 |
2.2 |
|
Somalia |
4.0 |
10.2 |
16.5 |
|
Ethiopia |
26.1 |
23.3 |
17.2 |
|
Sudan |
18.8 |
18.1 |
12.8 |
|
Total |
78.1 (147)a |
62.0 (103) |
59.1 (125) |
|
Percentage of African total |
53% |
60% |
47% |
a Figures in brackets give the total number of animals in the 37 countries in Africa infested with tsetse. Source: FAO/WHO/OIE (1982).
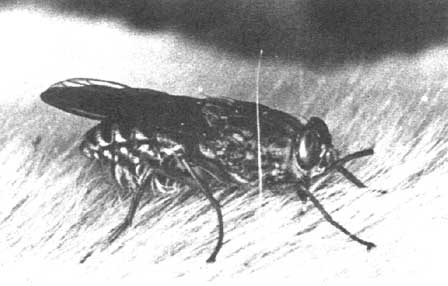
There is probably no other continent dominated by one disease to the same extent as is Africa by tsetse-transmitted trypanosomiasis. Many factors contribute to the magnitude of the problem, one of the major ones being the complexity of the disease itself. In cattle, three species of trypanosome, Trypanosoma congolense, T. vivax and T. brucei (Figure 3), cause the disease, either individually or jointly. These trypanosomes are transmitted cyclically by several different species of tsetse (genus Glossina) (Figure 4), each of which is adapted to different climatic and ecological conditions (Ford, 1971). While tsetse are not the only vectors of African trypanosomes, cyclical transmission of infection represents the most important problem, because once the tsetse fly becomes infected it remains infective for a long period, in contrast to the ephemeral nature of non-cyclical transmission. At the same time, trypanosomes infect a wide range of hosts including wild and domestic animals. The former, particularly the wild Bovidae and Suidae, do not suffer severe clinical disease but become carriers and constitute an important reservoir of infection (Murray et al, 1982). The success of the trypanosome as a parasite is to a large extent due to its ability to undergo antigenic variation, i.e. to change a single glycoprotein (Cross, 1975) which covers the pellicular surface, thereby enabling evasion of host immune responses and the establishment of persistent infections. Added to the complexity of multiple variable antigen types expressed during a single infection, each trypanosome species comprises an unknown number of different strains, all capable of elaborating a different repertoire of variable antigen types (Van Meirvenne et al, 1977).
For these reasons, no vaccine is available for use in the field, and the current control measures, which involve tsetse control or the use of trypanocidal drugs, have been limited in their efficacy.
Figure 3. African trypanosomes.
However, recent developments in the understanding of tsetse biology, antigenic variation, host susceptibility and immune responses, and in the strategic use of drugs currently available, means that all these approaches must be reevaluated with a view to improving the control of trypanosomiasis in domestic livestock.
Tsetse control
Future prospects for vaccination
Trypanotolerance
Trypanocidal drugs
Attempts to control tsetse have been made for over 60 years. Initially, they included eradication of wildlife, clearing of fly barriers to prevent the advance of the vector and widespread bush clearing to destroy breeding habitats. Following the introduction of modern chemicals, the principal method employed to control tsetse populations has been the use of insecticides, alone or in conjunction with traps and screens. Biological control methods are still under development and consideration.
Insecticides. The use of insecticides is the major method currently employed for tsetse control. The insecticides used fall into two categories, residual and non-residual. Residual insecticides (DDT and more recently dieldrin) are usually applied using hand-operated sprays that deliver the insecticide to sites where resting tsetse are known to alight. Ideally, the persistence of the insecticide should be sufficient to make only one application necessary. Non-residual insecticides require several applications; at present, endosulphan is the insecticide most frequently chosen. However, new synthetic pyrethroids which are 50 times more toxic to tsetse than endosulphan are currently being developed and tested. Non-residual treatments are applied mainly by fixed-wing aircraft or helicopters. Recent advances in the techniques of aerial spraying have resulted in better use of sprayed insecticides, by using fixed-wing aircraft against savanna species of tsetse and helicopters against tsetse inhabiting gallery forests and riverine areas. There has been significant success where insecticide control measures are properly implemented. For example, using residual sprays applied by hand-spraying machines nearly 200000 km2 of Nigeria have been cleared of Glossina morsitans. Control of tsetse by insecticides has also achieved notable success in Zimbabwe, Botswana and Zambia.
Despite the proven efficacy of tsetse control by insecticides, major tsetse advances are occurring in West, central, East and southern Africa. This is due to the severe limitations of this approach on practical, economic and environmental grounds. Currently in Africa, there is a lack of trained personnel, both at the leadership and field levels, to implement insecticide control programmes. The costs of insecticide control programmes are high, being approximately US$ 800, US$ 400, and US$ 2000/km2 for ground, fixed-wing aircraft and helicopter spraying respectively. Natural or man-made barriers are required to defend sprayed areas and prevent reinvasion, and constant surveillance for early detection of reinvasion is essential. Finally there are increasing demands to limit the use of insecticides because of their detrimental effect on naturally occurring fauna and flora.
Current research on insecticides involves the development of new potent chemicals with low toxic environmental effects e.g. synthetic pyrethroids such as permethrin, cypermethrin and decamethrin. In addition, the techniques of aerial spraying are being improved through studies of droplet size, rate of delivery and extent of dispersion.
Use of traps and screens. Traps and screens have been used for many years as means of sampling tsetse populations. However, with recent developments in the design and colour of traps and with the identification of tsetse attractants, increasing interest is being given to the use of traps as a possible method of tsetse control. It has been found that bovine breath acts as a powerful attractant for tsetse, with carbon dioxide and acetone being its major components (Vale, 1980). Colour is also an important attractant with responses of tsetse to yellow, green and blue being in the ratio of 1 to 10 to 150 respectively. Insecticide-impregnated traps have produced good results on a limited scale in Burkina Faso and Ivory Coast as a means of tsetse control in areas where human sleeping sickness is common. In Zimbabwe, it has been shown that the use of traps in combination with attractants leads to the capture of large numbers of G. m. morsitans and G. pallidipes. The use of improved traps impregnated with insecticide could develop into a simple and relatively cheap method of control, although it must be emphasised that this approach has yet to be proved effective in the field or with species of tsetse other than G. m. morsitans and G. pallidipes.
Biological methods. The concept of 'sterile male release' is based on the fact that tsetse females copulate only once, and if the male of a copulating pair is sterile the female will not produce during her lifetime. Thus, a series of field trials were carried out to evaluate the impact of the release of g -irradiated male tsetse on tsetse populations. Trial work was carried out at Mkwaja Ranch in Tanzania and in Zimbabwe, Burkina Faso, Nigeria and Zambia. Although under field conditions it has been demonstrated that this approach can significantly reduce tsetse populations, the majority opinion is that sterile male release is not a practical proposition because it is too sophisticated and too expensive. It is also estimated that ten sterile males are required per female. In order to reduce the number of sterile males required it is necessary to carry out two to three insecticide sprays and then to release 12000 sterile males per km2, even in areas where the tsetse density is low. The cost of this method is estimated to be US$ 3000 to US$ 4000 per km2. As the effect is tsetse-species specific, the cost increases with the number of species to be controlled.
Variation in susceptibility to trypanosome infection has been reported with certain strains of tsetse and consideration is being given to the development of tsetse which are refractory to infection. Research is also being carried out on the isolation of pheromones (sex hormones) and their characterisation for use as attractants in traps. Insect growth regulators have been identified with a view to their use as biological insecticides which would have no undesirable side-effects on humans or their environment. Enzymes in tsetse saliva and gut are also being identified with a view to using them as immunogens to vaccinate the host against tsetse. A further approach is the search for predators, parasites or pathogens which might serve to control tsetse populations. All these investigations are at a very early stage of development and much has to be done before they can be considered as possible methods of control.
There is no evidence that immune responses against antigens common to trypanosomes contribute to host protection. Thus, the major constraint to the development of a commercially applicable vaccine against trypanosomiasis is the phenomenon of antigenic variation, as it is now well established that host protective responses are effected by antibodies directed against surface coat antigens of the trypanosome (Murray and Urquhart, 1977). The repertoire of these antigens generated by bloodstream forms of the parasite is large, with the result that the development of a field vaccine has been considered an impossibility.
However, there is now evidence to indicate that the repertoire of antigens produced by metacyclic parasites following transmission through the tsetse is much more limited (Crowe et al, 1983). Thus, it has been possible to immunise cattle against tsetse-transmitted homologous (but not heterologous) strains by prior exposure to metacyclic parasites which have been propagated in tissue culture, or by prior infection via tsetse flies followed by trypanocidal drug treatment (Morrison et al, in press). The immunity produced has been shown to last for as long as 5 months. Nevertheless, the feasibility of production and the efficacy of a vaccine against metacyclic trypanosomes will depend on the relative stability of the metacyclic antigen repertoire for each species of trypanosome and on the number of strains which occur in the field. Current research is directed towards these objectives. It is thought, however, that the number of strains of T. congolense, T. vivax and T. brucei is likely to be prohibitively large for the production of a 'cocktail' vaccine containing the appropriate metacyclic antigens. As a result the development of a conventional vaccine against African trypanosomiasis is unlikely in the foreseeable future.
It has long been recognised that certain breeds of cattle in West and central Africa, mainly the N'Dama and West African Shorthorn, along with their sheep (Djallonke) and goat (Dwarf West African) counterparts, possess the ability to survive and be productive in tsetse-infested areas where other breeds rapidly succumb to trypanosomiasis (ILCA, 1979; Murray et al, 1982). However, it was only recently that the potential of these breeds was fully appreciated when it was established that they are much more productive than originally thought (ILCA, 1979) and that their trypanotolerance is an innate and not merely an acquired characteristic (Murray et al, 1982). As a result, increasing attention is being paid to the more widespread use of trypanotolerant breeds, especially in the tsetse-infested humid and subhumid areas of West and central Africa.
One of the major constraints to the more widespread use of trypanotolerant breeds of cattle is the limited number of animals available. At present, there are about 3.4 million N'Dama and 1.8 million West African Shorthorn, with the result that the current demand for N'Dama heifers and bulls greatly exceeds the number available for distribution. Furthermore, the cost of transport over great distances, along with the period of acclimatisation required for adaptation to different environmental conditions, means that a long-term investment is needed before any significant development and economic returns can be expected.
It must also be emphasised that the degree of trypanotolerance can be influenced by a number of factors, one of the most important being the level of tsetse challenge. It has been shown that as the level of challenge increases, productivity falls (Table 3) and that when tsetse challenge is high N'Dama can suffer severely from trypanosomiasis as judged by stunting, wasting, abortion, extreme lethargy leading to reduce ability to work and even death.
Thus, trypanotolerance is not a refractory state and it is essential in tsetse-infested areas to consider the strategic use of trypanocidal drugs in order to realise the full potential even of trypanotolerant breeds.
Table 3. Influence of level of tsetse challenge on productivity of trypanotolerant cattle, West and central Africa, 1977-78.
|
Level of tsetse challenge |
No. of herds |
Productivity indexa (kg) |
|
Zero |
3 |
40.1 |
|
Low |
13 |
31.9 |
|
Medium |
10 |
23.2 |
|
High |
4 |
18.8 |
a Total weight of 1-year-old calf and liveweight equivalent of milk produced per 100 kg of cow per year.
Source: ILCA (1979)
Escalating costs and other problems (discussed above) of initiating and maintaining tsetse control campaigns, together with the non-availability of a vaccine, have led to the livestock industries in the vast tsetse-infested areas of Africa being almost completely reliant on the use of trypanocidal drugs to both treat and prevent the disease. Without these drugs, the situation would be disastrous. However, despite the need and demand for effective trypanocides, no new drug has been produced for commercial use in the last 25 years1. As far as the treatment of cattle is concerned, only "Samorin" (isometamidium chloride - prophylactic), "Berenil" (diminazene aceturate-therapeutic) and "Novidium" or "Ethidium" (homidium chloride or bromide - therapeutic) are commercially available and there would appear to be no immediate prospects for new compounds for commercial use. None of the new compounds which have been identified in recent years has been found to be as effective in practice as those currently available. In fact over the last 10 years there have been increasing reports of the successful use of Samorin as a prophylactic, in combination with Berenil as a therapeutic.
1 Recently quinapyramine sulphate and prosalt have been reintroduced; however the special advantage of quinapyramine salts is against infections due to T. evansi.
Samorin has been successfully employed to control trypanosomiasis in exotic cattle in Kenya (Mwongela et al, 1981) and Zebu cattle in Kenya (Wilson et al, 1975), Tanzania (Blaser et al, 1979; Wiesenhutter et al, 1968), Ethiopia (Bourn and Scott, 1978) and Mali (Logan et al, 1984). In Mali, it was concluded that the economic losses caused by death and decreased weight gain in cattle treated with Berenil indicated that even in areas where the tsetse challenge was marginal and the incidence of trypanosomiasis relatively low, it paid to use a prophylactic programme with isometamidium chloride. Despite this considerable body of experimental evidence, the amount of trypanocidal drugs used in Africa is small in relation to the number of animals at risk. Currently, the number of doses employed is around 25 million (Le Roux, pers. comm.). This is despite the fact that a possible 50 million cattle, 30 million sheep and 40 million goats are estimated to be exposed. Even if animals were treated only twice a year, 240 million doses would be required, ten times the number currently used. The reasons for failure to make better use of the trypanocidal drugs available are complex but include:
a. Lack of precise information on the importance of the impact of trypanosomiasis on livestock production. Too often, widespread morbidity and mortality due to trypanosomiasis are attributed to poor nutrition. As a result, insufficient funds are allocated by governments for the purchase of trypanocidal drugs.b. The belief that the cost of trypanocidal drugs and of their use is high. This is a reasonable but probably unfounded assumption as there is little published information on the economics of use of trypanocidal drugs on livestock production.
c. The belief that repeated use of the same trypanocidal drugs must lead to drug resistance. This conclusion is supported by several field reports of the development of parasitaemia within a few weeks of treatment, an observation usually taken as evidence of drug resistance. While this may be true, it is equally likely to be the result of a relapse from a privileged site in the host to which the drug has no access, or to reinfection, or to underdosing. Weigh scales, and even weighbands, are frequently not available to allow accurate dose computation. Undoubtedly, true drug resistance does occur in terms of reduced parasite sensitivity to the drug (Pinder and Authie, 1984) and it must be given serious consideration where extensive use of trypanocidal drugs is being made or considered. However, to date there is little evidence that drug resistance is a problem in situations where management of the drug regime is good, even when the drug has been used over long periods of time.
In conclusion, the cost of tsetse control, the lack of a field vaccine and the limited prospects of new families of trypanocidal drugs appearing in the next 5 years make reliance on the trypanocidal drugs currently available an unavoidable necessity. It is therefore essential, firstly that their effectiveness be clearly demonstrated under different systems of management and various levels of tsetse challenge, and secondly that the risk of drug resistance be reduced.
It has been noted by a number of authors that the Mkwaja Ranch in Tanzania has over many years built up a collection of records on animal productivity and disease, probably unique in Africa. In early 1982, discussions were held at Mkwaja between representatives of May and Baker Ltd., the International Livestock Centre for Africa (ILCA), the International Laboratory for Research on Animal Diseases (ILRAD), and staff of Amboni Ltd. It was concluded that a major, well designed study of matching productivity and health data, linked to previous ecological and tsetse control work, would be of outstanding value. This would provide the opportunity to evaluate on a large scale the effectiveness of the use of Samorin (isometamidium chloride) as a prophylactic on grade Boran cattle exposed to a severe tsetse challenge.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}