![]()
![]()
![]()
Age and weight at early parturitions
Litter size, parturition interval and annual reproductive rate
Effects of climate on the period of birth and on litter size
Numbers of births by age class
Discussion
Reproductive performance has been analysed under three main headings: early reproductive performance; overall reproductive performance including litter size, parturition intervals and annual reproductive rate; and effects of a number of climatic variables on reproduction. Most data result from the long-term study although some data from specific short-term studies have been included where considered appropriate.
Histograms of the observed distributions of weight at first conception are given in Figure 40 for both goats and sheep. The mean weights (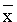 ± s.d.) for goats were 16.6 ± 3.46 kg in the range 6.8 - 29.5 kg and for sheep 22.8 ± 4.29 kg in the range 12.8 - 45.6 kg.
± s.d.) for goats were 16.6 ± 3.46 kg in the range 6.8 - 29.5 kg and for sheep 22.8 ± 4.29 kg in the range 12.8 - 45.6 kg.
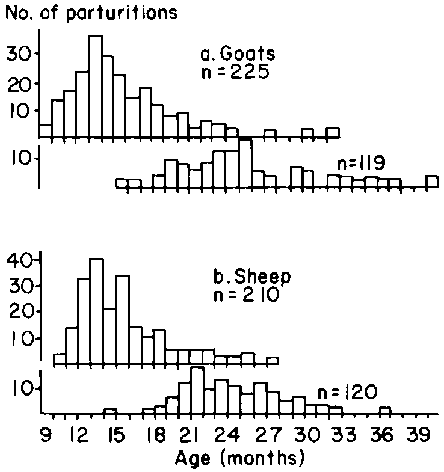
At first parturition the age of goats averaged 485 ± 128.87 days in the range 275 - 1104 days while for sheep age at first parturition was 480.2 ± 115.27 days. At second parturition the figures for goats and sheep respectively were 760.7 ± 144.46 days in the range 471 - 1300 days and 756.3 ± 128.45 days in the range 546 - 1221 days. The frequencies of the observed distributions for these data are shown in Figure 41.
Figure 40. Distribution of weights at first conception for goats and sheer in central Mali.
Figure 41. Ages at first and second parturitions for goats and sheep in central Mali.
The mean-squares values for the three traits are laid out in Table 32 while the least-squares means are given in Table 33. There was a significant effect of system on weight at first conception, with animals from the rice area being significantly heavier (P<0.05 for goats and P<0.001 for sheep) than those from the millet subsystem. Month of birth and maternal parity also had a highly significant effect on sheep weights. Sheep weighed significantly (P<0.001) more than goats at first conception. System was the only variable to have a significant effect on age at first and second parturitions, with the effect being confined to sheep. Although in goats the system effect was not significant, those reared in the rice areas also had their first and second parturitions earlier. Flock within system had significant effects on all the traits, with the effects being much more marked in the naturally less well endowed millet area.
Table 32. Analysis of variance of weight at first conception and ages at first and second parturitions for goats and sheer in central Mali.
|
Source of variation |
Weight at first conception |
Age at first parturition |
Age at second parturition |
|||||||||
|
Goats |
Sheep |
Goats |
Sheep |
Goats |
Sheep |
|||||||
|
d.f. |
MS |
d.f. |
MS |
d.f. |
MS |
d.f. |
MS |
d.f. |
MS |
d.f. |
MS |
|
|
System |
1 |
429* |
1 |
17 978*** |
1 |
11 399 |
1 |
145 185*** |
1 |
8 516 |
1 |
101 710** |
|
Month of birth |
11 |
84 |
11 |
4 766*** |
11 |
16 998 |
11 |
15 121 |
11 |
20 496 |
11 |
9 796 |
|
Type of birth |
1 |
156 |
1 |
52 |
1 |
37 552 |
1 |
478 |
1 |
2 440 |
1 |
34 752 |
|
Parity |
7 |
35 |
7 |
3 976** |
7 |
18 006 |
7 |
12 901 |
7 |
8 640 |
7 |
14 160 |
|
Flock/millet |
12 |
357*** |
12 |
5 049*** |
12 |
39 217** |
12 |
30 442*** |
12 |
64 618*** |
12 |
18 934 |
|
Flock/rice |
4 |
130 |
5 |
320 |
4 |
90 408*** |
5 |
11,041 |
4 |
35 296 |
5 |
37 053* |
|
Error |
264 |
110 |
239 |
1 324 |
270 |
14 649 |
242 |
10 269 |
129 |
16 135 |
133 |
13 201 |
***P<0.001; **P<0.01; *P<0.05.
Table 33. Least-squares meant for weight at first conception and ages at first and second parturitions for goats and sheep in central Mali.
|
Variable |
Weight (kg) at first conception |
Age (days) at first parturition |
Age (days) at second parturition |
||||||||||
|
Goats |
Sheep |
Goats |
Sheep |
Goats |
Sheep |
||||||||
|
n |
|
n |
|
n |
|
n |
|
n |
|
n |
|
||
|
Raw mean |
301 |
16.6 |
277 |
22.8 |
307 |
485.5 |
280 |
480.2 |
166 |
760.7 |
171 |
756.3 |
|
|
Overall LS mean |
301 |
17.1 |
277 |
22.8 |
307 |
496.9 |
280 |
463.9 |
166 |
762.8 |
171 |
721.8 |
|
|
System |
|||||||||||||
|
|
Millet |
239 |
16.4a |
186 |
21.6a |
244 |
508.2 |
189 |
497.0a |
136 |
779.8 |
121 |
760.0a |
|
|
Rice |
62 |
17.8b |
91 |
24.0b |
63 |
485.6 |
91 |
430.7b |
30 |
745.9 |
50 |
683.6b |
|
Month of birth |
|||||||||||||
|
|
January |
13 |
17.3 |
30 |
21.2ad |
14 |
467.8 |
30 |
424.9 |
10 |
785.6 |
12 |
676.3 |
|
|
February |
17 |
15.4 |
17 |
23.0ac |
17 |
363.1 |
17 |
480.8 |
14 |
674.9 |
9 |
699.5 |
|
|
March |
33 |
17.5 |
26 |
22.2aef |
34 |
490.2 |
26 |
472.6 |
22 |
765.5 |
21 |
731.7 |
|
|
April |
24 |
18.1 |
16 |
21.3agh |
24 |
554.8 |
16 |
481.5 |
14 |
793.1 |
11 |
764.7 |
|
|
May |
15 |
17.0 |
15 |
23.0aij |
16 |
468.1 |
16 |
479.3 |
5 |
680.1 |
11 |
711.7 |
|
|
June |
12 |
17.3 |
16 |
24.1bceglkin |
12 |
543.7 |
16 |
499.4 |
5 |
778.5 |
11 |
763.8 |
|
|
July |
10 |
17.6 |
19 |
22.9akm |
10 |
529.3 |
19 |
457.4 |
4 |
872.1 |
9 |
750.3 |
|
|
August |
8 |
16.7 |
15 |
26.6n |
8 |
479.4 |
15 |
509.0 |
2 |
834.5 |
9 |
755.1 |
|
|
September |
45 |
17.3 |
35 |
23.9bcjmo |
45 |
495.3 |
35 |
460.1 |
29 |
770.5 |
22 |
705.9 |
|
|
October |
37 |
17.3 |
28 |
22.6alo |
40 |
494.0 |
28 |
426.2 |
23 |
709.4 |
17 |
716.8 |
|
|
November |
53 |
16.6 |
32 |
20.7dfhp |
53 |
510.1 |
32 |
444.2 |
15 |
762.5 |
20 |
684.8 |
|
|
December |
34 |
16.7 |
28 |
21.5ap |
34 |
466.8 |
30 |
431.0 |
13 |
727.3 |
19 |
700.9 |
|
Type of birth |
|||||||||||||
|
|
Single |
193 |
17.4 |
260 |
22.9 |
198 |
483.5 |
262 |
460.7 |
105 |
768.0 |
156 |
753.9 |
|
|
Twin/triplet |
108 |
16.8 |
17 |
22.7 |
109 |
510.3 |
18 |
467.0 |
61 |
757.7 |
15 |
689.6 |
|
Parity |
|||||||||||||
|
|
'0' |
124 |
16.8 |
58 |
25.1a |
129 |
492.9 |
60 |
497.2 |
79 |
791.3 |
39 |
749.4 |
|
|
1 |
46 |
16.8 |
58 |
23.0bc |
46 |
517.7 |
58 |
486.7 |
26 |
760.3 |
32 |
757.9 |
|
|
2 |
43 |
16.6 |
62 |
23.8acde |
43 |
480.0 |
62 |
479.5 |
25 |
757.6 |
35 |
707.3 |
|
|
3 |
33 |
16.9 |
46 |
22.1bf |
33 |
523.6 |
47 |
444.6 |
15 |
791.1 |
33 |
710.6 |
|
|
4 |
23 |
17.8 |
24 |
22.4bdg |
23 |
501.4 |
24 |
464.6 |
11 |
782.2 |
16 |
719.0 |
|
|
5 |
15 |
16.7 |
13 |
22.7acfgh |
16 |
489.3 |
13 |
469.3 |
8 |
754.2 |
7 |
772.4 |
|
|
6 |
10 |
17.2 |
8 |
21.9beh |
10 |
414.5 |
8 |
430.9 |
1 |
592.4 |
5 |
645.7 |
|
|
7-10 |
7 |
17.7 |
8 |
21.1beh |
7 |
555.8 |
8 |
438.0 |
1 |
873.7 |
4 |
712.1 |
Note: Parity '0' = all unknown parities assumed to be ³ 4.Within variable groups, means followed by different letters differ significantly (P<0.05). Variable groups without any letters did not show a significant difference in the analysis of variance.
There was no significant correlation between age at first parturition and the interval to the second parturition either for goats (r = 0.20, P>0.05) or for sheep (r = 0.03, P>0.05).
Annual reproductive rate (ARR) was calculated as a function of litter size and parturition interval according to the formula: litter size x 365/subsequent parturition interval. Calculations for parturition interval and ARR were limited to animals having given a previous birth in 1983 in order that short intervals would not bias the results.
During the study period there was a total of 3605 parturitions giving rise to 4049 young. The distribution of births and young is shown in Table 34. Mean litter size ( ± s.d.) for goats was 1.19 ± 0.41 and for sheep 1.04 ± 0.21. Litter size increased from 1.04 for primiparous goats to a maximum of 1.39. In sheep, litter size varied from 1.01 for primiparous females to 1.13 in older animals.
± s.d.) for goats was 1.19 ± 0.41 and for sheep 1.04 ± 0.21. Litter size increased from 1.04 for primiparous goats to a maximum of 1.39. In sheep, litter size varied from 1.01 for primiparous females to 1.13 in older animals.
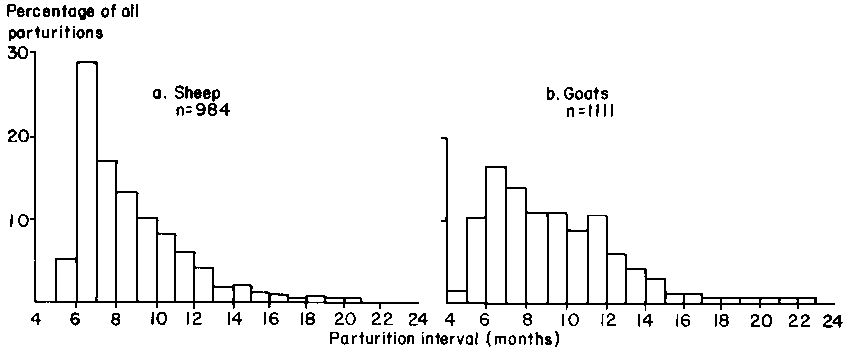
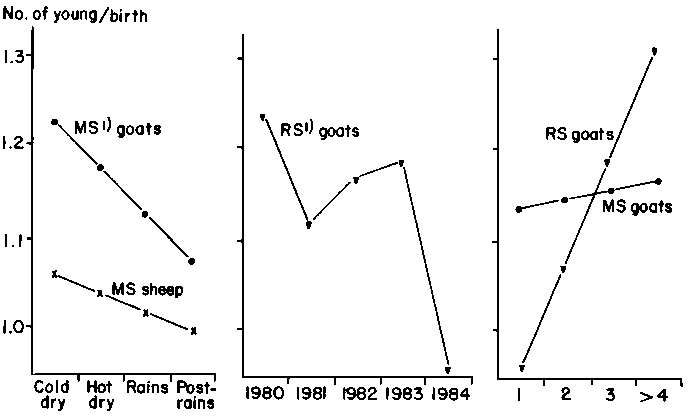
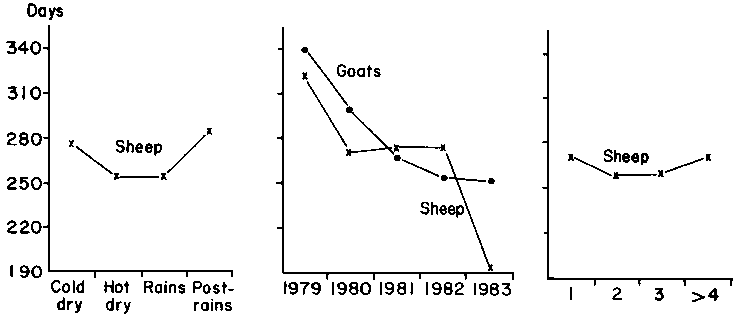
The observed distribution of parturition intervals is shown in Figure 42. The mean for goats was 291 ± 105.2 days and for sheep 261 ± 76.3 days. The longest interval for goats (298 ± 99.2 days) occurred between the first and second parities and the shortest (208 ± 45.4 days) after the ninth parity. For sheep, the longest interval (208 ± 63.6 days) was again after the first parity, with a decrease in the length of the interval in subsequent parities. Overall annual reproductive rates, calculated from the two previous parameters, were 1.49 for goats and 1.45 for sheep.
These data from the long-term study can be compared with those from a rapid survey carried out in early 1978 which attempted to establish the distribution of types of birth and litter size by age class. The main results from this small study are shown in Table 35.
Mature animals with an unknown breeding history were not included in the least-squares analyses. Parity in this set of analyses refers to the order of births in the reproductive career of the animal whose records are being analysed and not, as in the section on early reproductive performance, to the parity of its dam. Parities greater than 4 were grouped and treated as fourth parity and triplet and twin parturitions were grouped as multiple births. A separate analysis was carried out for each parameter for each species. The models considered the random effect of dam within system and within flock and the fixed effects of system, flock, season and year of parturition, type of birth, sex of offspring and parity. During preliminary analyses, system (rainfed millet and irrigated rice) was seen to have significant effects on some of the traits being studied. Because of the limitations of the models, the main analyses were therefore carried out by systems with dams within flocks as the random variable. Orthogonal polynomials were fitted to some of the main variables in an attempt to further understand some of the influences acting on them.
Levels of significance for the different sources of variation acting on the three measured traits are shown in Table 36. System influenced significantly all three traits in sheep but none at all in goats. All further analyses resulting from this analysis of variance were however carried out by system with flock introduced as a fixed effect. The practical results of the effects of system on sheep were that those in the rice subsystem had a better total reproductive performance than those in the millet subsystem. This was probably due to the better year-round feed conditions in the former subsystem, a hypothesis supported by the lack of significance in the seasonal effect on all three traits for sheep.
Table 34. Numbers of parturitions and percentages of young by parturition type for sheep and goats in central Mali.
|
Type of birth |
Sheep |
Goats |
||
|
No. |
% |
No. |
% |
|
|
Total births |
1 650 |
100.0 |
1 955 |
100.0 |
|
Total young |
1 722 |
100.0 |
2 327 |
100.0 |
|
Single births |
1 579 |
95.7 |
1 593 |
81.5 |
|
Young born as singles |
1 579 |
91.7 |
1 593 |
68.5 |
|
Twin births |
70 |
4.2 |
350 |
17.9 |
|
Young born as twins |
140 |
8.1 |
700 |
30.1 |
|
Triplet births |
1 |
0.1 |
12 |
0.6 |
|
Young born as triplets |
3 |
0.2 |
36 |
1.5 |
Figure 42. Frequency distribution of parturition intervals for sheep and goats in central Mali.
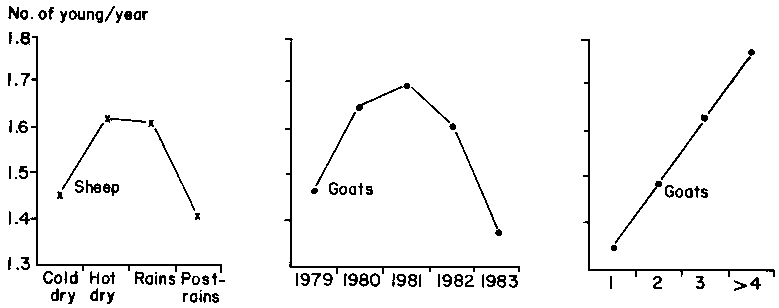
The least-squares means for the three traits are shown in Table 37 for sheep and in Table 38 for goats. Table 39 provides data on the best-fit polynomials for variables where significance occurred in the main analyses, and Figure 43 shows in graphic form the calculated orthogonal fits. Other than the effects of the flock factor, and with the exception of the effects of year and parity on litter size in goats in the rice subsystem, all significant effects are confined to one or the other species in the millet subsystem.
For these analyses the numbers of births and the mean number of young per birth were calculated for each 10-day period from 1978 to 1983. Simple correlations and multiple linear regressions were then calculated between these data and six climatic variables for each of the 15th to 20th 10-day periods prior to those being analysed for numbers of parturitions and litter size. BMDP programmes (Dixon and Brown, 1983) were used for the statistical analyses.
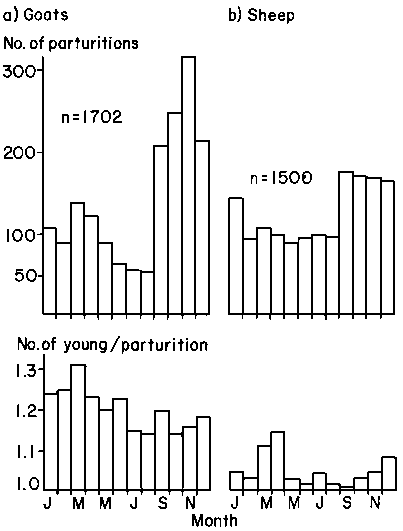
The distribution of 1702 parturitions for goats and 1500 for sheep by month is shown in Figure 44 which also shows the mean litter size by month. There were highly significant differences among months for the numbers of parturitions in goats (X² = 553.8, d.f. = 11, P<0.001) and for those in sheep (X² = 108.0, d.f. = 11, P<0.001). The minimum number of parturitions (54) in goats occurred in August and the maximum (323) in November. In sheep, the minimum number (88) was in May, the maximum (174) being in September.
Litter size in goats varied from a maximum of 1.31 kids in March to a minimum of 1.14 in both August and October. In sheep, maximum litter size of 1.14 lambs was achieved in April with a minimum of 1.01 in September. An analysis of variance showed significant differences among months for both goats (F = 5.55, d.f. = 11,1657, P<0.001) and sheep (F = 3.90, d.f. = 11,1444, P<0.001).
The maximum number of conceptions in both goats and sheep took place in May to July while the maximum number of ova appeared to be shed in October and November in goats and perhaps slightly later (in November and December) in sheep. The smallest number of conceptions in goats took place in February and March, while in sheep there was a fairly clearly defined period of poor conception rates extending from October through March. Monitoring actual oestrus activity in sheep in the Egyptian subtropics, Aboul-Naga et al (1985) found it to be irregular throughout the year but with a minimum in April. Monitored by hormone levels in Niger, in climatic conditions similar to those at Niono, minimum oestrus activity and longest cycles were found from January to April (Yenikoye et al, 1982). In the latter case, abnormally long cycles contributed to the low numbers of conceptions observed: there was, however, a rapid upsurge in oestrus activity from May onwards.
Correlations between goat parturitions and climatic variables were positive and highly significant for maximum temperature (r = 0.30 to 0.47), minimum temperature (r=0.25 to 0.42) and day length (r = 0.27 to 0.40) for all of the 10-day periods from the 15th to the 20th previously. A similar pattern was observed for sheep with correlations ranging from 0.29 to 0.39 for maximum temperature, from 0.17 to 0.37 for minimum temperature, and from 0.25 to 0.42 for day length. There were significant positive correlations also between the parturition interval and both maximum and minimum humidity at the 15th period for both goats and sheep and between parturition interval and rainfall at the 15th period for sheep, but no other periodic conditions were significant for these parameters and some were negative.
Table 35. Reproductive performance of agropastoral goats and sheep, based on owners' recall data.
|
Sheep |
Goats |
||||||||||||
|
4 pairs |
3 pairs |
2 pairs |
1 pair |
Temporary |
Overall |
4 pairs |
3 pairs |
2 pairs |
1 pair |
Temporary |
Overall |
||
|
Number in sample |
37 |
21 |
22 |
42 |
16 |
138 |
180 |
77 |
61 |
111 |
15 |
444 |
|
|
Type of birth |
|||||||||||||
|
|
Single |
124 |
52 |
43 |
41 |
7 |
267 |
457 |
139 |
81 |
77 |
5 |
759 |
|
|
Twin |
9 |
2 |
2 |
1 |
|
14 |
203 |
11 |
4 |
1 |
|
219 |
|
|
Triplet |
1 |
|
|
|
|
1 |
8 |
|
|
|
|
8 |
|
Total parturitions |
134 |
54 |
45 |
42 |
7 |
282 |
668 |
150 |
85 |
78 |
5 |
986 |
|
|
Total young born |
145 |
56 |
47 |
43 |
7 |
298 |
887 |
161 |
89 |
79 |
5 |
1 221 |
|
|
Average litter size |
1.08 |
1.04 |
1.04 |
1.02 |
1.00 |
1.06 |
1.33 |
1.07 |
1.05 |
1.01 |
1.00 |
1.24 |
|
Table 36. Analysis of variance of litter size, parturition interval and annual reproductive rate for sheep and goats in central Mali.
|
Trait/species |
Source of variation |
||||||||
|
System |
Flock |
Dam/Flock |
Season |
Year |
Parity |
Sex of young |
Type of birth |
||
|
Litter size |
|||||||||
|
|
All sheep |
** |
- |
n.s. |
** |
n.s. |
n.s. |
- |
- |
|
|
MSa sheep |
- |
*** |
n.s. |
** |
n.s. |
n.s. |
- |
- |
|
|
RSa sheep |
- |
n.s. |
n.s. |
n.s. |
n.s. |
n.s. |
- |
- |
|
|
All goats |
n.s. |
- |
n.s. |
** |
n.s. |
*** |
- |
- |
|
|
MS goats |
- |
** |
n.s. |
*** |
n.s. |
*** |
- |
- |
|
|
RS goats |
- |
*** |
n.s. |
n.s. |
* |
* |
- |
- |
|
Parturition interval |
|||||||||
|
|
All sheep |
*** |
- |
** |
** |
n.s. |
* |
* |
n.s. |
|
|
MS sheep |
- |
*** |
* |
*** |
** |
n.s. |
* |
n.s. |
|
|
RS sheep |
- |
** |
* |
n.s. |
n.s.. |
n.s. |
n.s. |
n.s. |
|
|
All goats |
n.s. |
- |
** |
* |
** |
n.s. |
n.s. |
n.s. |
|
|
MS goats |
- |
*** |
** |
n.s. |
* |
n.s. |
n.s. |
n.s. |
|
|
RS goats |
- |
n.s. |
** |
n.s. |
n.s. |
n.s. |
n.s. |
n.s. |
|
Annual reproductive rate |
|||||||||
|
|
All sheep |
*** |
- |
** |
** |
n.s. |
n.s. |
n.s. |
- |
|
|
MS sheep |
- |
*** |
n.s. |
** |
n.s. |
n.s. |
n.s. |
- |
|
|
RS sheep |
- |
* |
** |
n.s. |
n.s. |
n.s. |
n.s. |
- |
|
|
All goats |
n.s. |
- |
n.s. |
n.s. |
* |
* |
n.s. |
- |
|
|
MS goats |
- |
** |
n.s. |
n.s. |
* |
* |
n.s. |
- |
|
|
RS goats |
- |
n.s. |
n.s. |
n.s. |
n.s. |
n.s. |
n.s. |
- |
***P<0.001; **P<0.01; *P<0.05; n.s. = not significant.
a MS = millet subsystem; RS = rice subsystem.
The litter size of goats showed significant correlations with maximum temperature (r = 0.14 to 0.19) at the 17th to 19th periods and with both maximum humidity (r = 0.16 to 0.22) and minimum humidity (r = 0.14 to 0.22) at the 17th to 20th periods previously. In sheep, litter size was significantly correlated only with maximum humidity (r = 0.16 to 0.26) and minimum humidity (r = 0.15 to 0.26) for all of the 15th to 20th periods in the case of the former and from the 17th period in the case of the latter.
When stepwise multiple regression was done, the final equations containing terms significant at the 5% level accounted for a low proportion of the variance. The maximum variance accounted for by parturitions in goats was 25.8% at the 17th period previously, with maximum temperature accounting for 19.9% of this and day length for the rest. For sheep, the maximum variance accounted for, at the 18th period, was 17.7% with maximum temperature contributing 14.6% and day length the rest.
The total amount of variance accounted for by litter size was much lower, being 6.7% at the 19th period with minimum humidity accounting for 3.7% and minimum temperature for 2.9%. In sheep, most variance (6.7% and 6.8% respectively) was accounted for by relative humidity at the 19th and 20th periods previously.
During the rapid study already referred to it was possible to obtain an indication of the numbers of births by each class of age of female. The main results are provided in Table 40. For sheep, ewes with milk teeth and one to four pairs of permanent incisors had given birth 0.43, 1.00, 2.05, 2.57 and 3.62 times respectively, the overall number of births per breeding female in the flock being 1.84. For goats, the numbers of births were 0.33, 0.70, 1.39, 1.95 and 3.71 for the individual age groups and 2.22 births for all does in the flock. For both species there was a rapid reduction in the number of females having given birth more than five times.
Early reproductive performance
Main reproductive traits
Climatic effects on reproduction
The mean age at first kidding of 463 days is in good agreement with the few other existing data for goats of a similar type managed under traditional systems in the West African semiarid zone. For example in Chad, it has been found that age at first parturition varies from 418 to 502 days depending on the region (Bertaudière, 1979; Dumas, 1980). In Niger, Gerbaldi (1978) found that Sahel-type goats first kid at 401 ± 24 days. In southern Niger, the Red Maradi goat had an age of 427 ± 13 days at first kidding (Haumesser, 1975).
The age at first lambing of 474 days also agrees well with other published reports. In Chad, mean age at first lambing varied from 395 to 502 days depending on the region (Gerbaldi, 1978; Haumesser and Gerbaldi, 1980).
Management practices to control breeding are generally designed to delay the age at first parturition. This applies whether the management is traditional, as in the case of the Maasai pastoral system (where an apron is used to cover the penis of the male and some flock stratification is evident) where the age at first kidding is 556 ± 119 days and at first lambing 549 ± 112 days (Wilson et al, 1984), or under station conditions, as in Senegal (where males are separated from the females when breeding is not desired and females are mated only on attaining a specified minimum weight) where age at first lambing is 572 ± 24 days (Fall et al, 1982).
The greater weight at first conception in the irrigated rice subsystem reflected the differences in food availability between the rice and millet subsystems, and the generally faster growth rates (from birth weights of 1.8 kg for goats and 2.6 kg for sheep in both subsystems) and heavier mature weights achieved in the rice subsystem (31.8 kg for goats and 35.9 kg for sheep), compared to the millet subsystem (28.1 and 32.1 kg respectively for goats and sheep). Earlier physical maturity in the rice subsystem at first conception also apparently results in slightly earlier physiological maturity, reflected in the 35-day younger age at first parturition.
The only other variable to affect significantly the age at first parturition is maternal parity, with offspring of young and Old mothers maturing later than those from dams in the intermediate (35 years) age groups. The reasons for this are not clear as there is no obvious relationship with weight at first conception.
Table 37. Least-squares means for litter size, parturition interval and annual reproductive rate of sheep in central Mali.
|
Variable |
Millet subsystem |
Rice subsystem |
|||||||||
|
Litter size |
Parturition interval |
Annual reproductive rate |
Litter size |
Parturition interval |
Annual reproductive rate |
||||||
|
n |
No. of young |
n |
Days |
Young/year |
n |
No. of young |
n |
Days |
Young/year |
||
|
Overall |
909 |
1.036 |
502 |
290 |
1.53 |
445 |
1.031 |
257 |
259 |
1.63 |
|
|
Flocka |
|||||||||||
|
|
1 |
85 |
1.012 |
40 |
304 |
1.37 |
86 |
1.032 |
31 |
257 |
1.59 |
|
|
2 |
35 |
1.021 |
19 |
251 |
1.69 |
160 |
1.089 |
102 |
237 |
1.85 |
|
|
3 |
39 |
1.152 |
26 |
262 |
1.99 |
74 |
1.045 |
49 |
228 |
1.73 |
|
|
4 |
50 |
1.022 |
27 |
317 |
1.40 |
58 |
0.984 |
36 |
241 |
1.65 |
|
|
5 |
207 |
1.011 |
115 |
265 |
1.61 |
24 |
1.006 |
13 |
310 |
1.37 |
|
|
6 |
101 |
1.016 |
65 |
271 |
1.56 |
43 |
1.033 |
26 |
279 |
1.57 |
|
|
7 |
54 |
1.016 |
28 |
308 |
1.40 |
- |
- |
- |
- |
- |
|
|
8 |
78 |
1.010 |
45 |
302 |
1.42 |
- |
- |
- |
- |
- |
|
|
9 |
31 |
1.117 |
20 |
264 |
1.80 |
- |
- |
- |
- |
- |
|
|
10 |
122 |
1.033 |
59 |
288 |
1.54 |
- |
- |
- |
- |
- |
|
|
11 |
32 |
1.002 |
15 |
365 |
1.18 |
- |
- |
- |
- |
- |
|
|
12 |
37 |
0.999 |
19 |
303 |
1.36 |
- |
- |
- |
- |
- |
|
|
13 |
38 |
1.053 |
24 |
275 |
1.57 |
- |
- |
- |
- |
- |
|
|
Average SE |
|
0.0236 |
|
18.1 |
0.096 |
|
0.0316 |
|
19.6 |
0.115 |
|
Season |
|||||||||||
|
|
Cold dry |
271 |
1.053 |
147 |
297 |
1.46 |
101 |
1.039 |
45 |
262 |
1.62 |
|
|
Hot dry |
206 |
1.074 |
117 |
285 |
1.63 |
143 |
1.054 |
90 |
256 |
1.74 |
|
|
Rains |
217 |
1.004 |
117 |
267 |
1.62 |
103 |
0.991 |
60 |
257 |
1.62 |
|
|
Post-rains |
215 |
1.018 |
121 |
312 |
1.40 |
98 |
1.042 |
62 |
260 |
1.52 |
|
|
Average SE |
|
0.0144 |
|
13.0 |
0.054 |
|
0.0327 |
|
16.0 |
0.086 |
|
Year |
|||||||||||
|
|
1978 |
90 |
1.019 |
- |
- |
- |
11 |
0.932 |
- |
- |
- |
|
|
1979 |
101 |
1.023 |
81 |
306 |
1.48 |
26 |
1.054 |
16 |
222 |
2.01 |
|
|
1980 |
159 |
1.020 |
110 |
298 |
1.44 |
70 |
1.062 |
44 |
275 |
1.55 |
|
|
1981 |
159 |
1.023 |
122 |
296 |
1.47 |
79 |
1.070 |
60 |
258 |
1.66 |
|
|
1982 |
144 |
1.042 |
105 |
300 |
1.50 |
105 |
1.022 |
82 |
264 |
1.56 |
|
|
1983 |
166 |
1.065 |
84 |
252 |
1.75 |
107 |
1.048 |
55 |
274 |
1.36 |
|
|
1984 |
90 |
1.057 |
- |
- |
- |
47 |
1.032 |
- |
- |
- |
|
|
Average SE |
|
0.0263 |
|
15.3 |
0.078 |
|
0.0751 |
|
24.8 |
0.165 |
|
Parity |
|||||||||||
|
|
1 |
271 |
1.034 |
141 |
292 |
1.51 |
145 |
1.012 |
84 |
271 |
1.47 |
|
|
2 |
212 |
1.055 |
115 |
287 |
1.59 |
105 |
0.992 |
60 |
257 |
1.57 |
|
|
3 |
155 |
1.026 |
89 |
277 |
1.59 |
80 |
1.038 |
46 |
248 |
1.69 |
|
|
³ 4 |
271 |
1.028 |
157 |
306 |
1.42 |
115 |
1.078 |
67 |
257 |
1.78 |
|
|
Average SE |
|
0.0177 |
|
14.2 |
0.067 |
|
0.0396 |
|
17.8 |
0.104 |
|
Sex |
|||||||||||
|
|
Female |
- |
- |
272 |
281 |
1.56 |
- |
- |
116 |
256 |
1.55 |
|
|
Male |
- |
- |
230 |
300 |
1.49 |
- |
- |
141 |
261 |
1.70 |
|
|
Average SE |
|
|
|
11.7 |
0.039 |
|
|
|
14.2 |
0.067 |
|
Type of birth |
|||||||||||
|
|
Single |
- |
- |
487 |
269 |
- |
- |
- |
242 |
253 |
- |
|
|
Multiple |
- |
- |
15 |
312 |
- |
- |
- |
15 |
264 |
- |
|
|
Average SE |
|
|
|
15.4 |
|
- |
- |
|
16.8 |
|
a Flocks in the millet and rice subsystems carrying the same number are not necessarily under the same ownership.
Table 38. Least-squares means for litter size, parturition interval and annual reproductive rate of goats in central Mali.
|
Variable |
Millet subsystem |
Rice subsystem |
|||||||||
|
Litter size |
Parturition interval |
Annual reproductive rate |
Litter size |
Parturition interval |
Annual reproductive rate |
||||||
|
n |
No. of young |
n |
Days |
Young/year |
n |
No. of young |
n |
Days |
Young/year |
||
|
Overall |
1104 |
1.154 |
597 |
298 |
1.53 |
310 |
1.177 |
166 |
297 |
1.63 |
|
|
Flocka |
|||||||||||
|
|
1 |
71 |
1.182 |
29 |
272 |
1.58 |
38 |
1.323 |
21 |
286 |
1.75 |
|
|
2 |
174 |
1.236 |
102 |
267 |
1.79 |
39 |
1.275 |
21 |
279 |
1.82 |
|
|
3 |
173 |
1.160 |
106 |
255 |
1.84 |
16 |
1.253 |
7 |
251 |
1.93 |
|
|
4 |
42 |
1.310 |
20 |
270 |
1.83 |
50 |
1.062 |
28 |
295 |
1.49 |
|
|
5 |
39 |
1.106 |
21 |
328 |
1.39 |
147 |
1.084 |
82 |
295 |
1.52 |
|
|
6 |
23 |
1.245 |
13 |
297 |
1.68 |
20 |
1.066 |
7 |
377 |
1.29 |
|
|
7 |
22 |
1.043 |
11 |
325 |
1.28 |
- |
- |
- |
- |
- |
|
|
8 |
41 |
1.160 |
15 |
293 |
1.73 |
- |
- |
- |
- |
- |
|
|
9 |
311 |
1.182 |
171 |
302 |
1.44 |
- |
- |
- |
- |
- |
|
|
10 |
32 |
1.049 |
17 |
280 |
1.52 |
- |
- |
- |
- |
- |
|
|
11 |
30 |
1.095 |
16 |
343 |
1.24 |
- |
- |
- |
- |
- |
|
|
12 |
60 |
1.050 |
35 |
334 |
1.31 |
- |
- |
- |
- |
- |
|
|
13 |
39 |
1.152 |
19 |
313 |
1.27 |
- |
- |
- |
- |
- |
|
|
14 |
47 |
1.178 |
22 |
293 |
1.52 |
- |
- |
- |
- |
- |
|
|
Average SE |
|
0.0534 |
|
21.7 |
0.132 |
|
0.0599 |
|
27.9 |
0.161 |
|
Season |
|||||||||||
|
|
Cold dry |
260 |
1.245 |
140 |
316 |
1.50 |
69 |
1.161 |
27 |
287 |
1.67 |
|
|
Hot dry |
252 |
1.186 |
146 |
291 |
1.62 |
75 |
1.190 |
40 |
296 |
1.52 |
|
|
Rains |
184 |
1.076 |
114 |
281 |
1.56 |
39 |
1.144 |
31 |
283 |
1.79 |
|
|
Post-rains |
408 |
1.107 |
197 |
305 |
1.44 |
127 |
1.214 |
68 |
322 |
1.55 |
|
|
Average SE |
|
0.0301 |
|
12.0 |
0.072 |
|
0.0543 |
|
21.9 |
0.134 |
|
Year |
|||||||||||
|
|
1978 |
61 |
1.164 |
- |
- |
- |
- |
- |
- |
- |
- |
|
|
1979 |
99 |
1.184 |
70 |
364 |
1.40 |
- |
- |
- |
- |
- |
|
|
1980 |
148 |
1.208 |
101 |
304 |
1.68 |
46 |
1.298 |
33 |
303 |
1.72 |
|
|
1981 |
211 |
1.184 |
153 |
283 |
1.69 |
66 |
1.126 |
48 |
301 |
1.49 |
|
|
1982 |
223 |
1.110 |
153 |
278 |
1.51 |
67 |
1.269 |
49 |
313 |
1.54 |
|
|
1983 |
196 |
1.081 |
120 |
261 |
1.38 |
68 |
1.188 |
36 |
272 |
1.77 |
|
|
1984 |
166 |
1.143 |
- |
- |
- |
63 |
1.004 |
- |
- |
- |
|
|
Average SE |
|
0.0579 |
|
17.7 |
0.112 |
|
0.0888 |
|
24.5 |
0.158 |
|
Parity |
|||||||||||
|
|
1 |
370 |
1.004 |
197 |
274 |
1.37 |
102 |
1.028 |
54 |
299 |
1.41 |
|
|
2 |
256 |
1.093 |
157 |
294 |
1.36 |
67 |
1.060 |
41 |
295 |
1.54 |
|
|
3 |
186 |
1.205 |
106 |
299 |
1.65 |
58 |
1.277 |
30 |
271 |
1.87 |
|
|
³ 4 |
292 |
1.311 |
137 |
326 |
1.74 |
86 |
1.343 |
41 |
324 |
1.71 |
|
|
Average SE |
|
0.0368 |
|
14.7 |
0.095 |
|
0.0664 |
|
24.8 |
0.161 |
|
Sex |
|||||||||||
|
|
Female |
- |
- |
304 |
295 |
1.55 |
- |
- |
81 |
303 |
1.60 |
|
|
Male |
- |
- |
293 |
301 |
1.51 |
- |
- |
85 |
291 |
1.67 |
|
|
Average SE |
|
|
|
9.9 |
0.052 |
|
|
|
18.6 |
0.095 |
|
Type of birth |
|||||||||||
|
|
Single |
- |
- |
545 |
304 |
- |
- |
- |
152 |
283 |
- |
|
|
Multiple |
- |
- |
52 |
293 |
- |
- |
- |
14 |
312 |
- |
|
|
Average SE |
|
|
|
11.5 |
|
|
|
|
21.5 |
|
Note: a Flocks in the millet and rice subsystems carrying the same number are not necessarily under the same ownership.
On the whole the effects of environmental factors on age at first parturition are small. This was also found to be the case in Senegal where neither year, nor month nor type of birth had any significant effect on age at first lambing (Fall et al, 1982). It would appear, therefore, that little is to be gained by attempting to control breeding seasons to optimise age at first parturition. Mortality of young born to primiparous females is almost always higher than that of those born to older females. Although delaying first parturition to an age when the female is nearer to its full physical development reduces the death rate to some extent, the slower growth rate of the young is reflected in the lower productivity of this class of dam when compared with that of older dams There is no evidence from traditional systems that early first parturitions result in subsequent diminished reproductive capacity and it is therefore probably worthwhile selecting for an early first parturition, accepting the higher death rate and lower productivity resulting from this and anticipating an extra parturition and a longer total reproductive life. Such a strategy might also enable a greater proportion of breeding females in the middle age groups to be maintained in relation to primiparous and older females, thus leading to further improvements in total flock productivity.
The effects of parity are those which might logically be expected. Litter size increased linearly with age in goats in both subsystems. The lack of significance in sheep is probably due to the overall low level of twinning. The quadratic form of the parturition interval of sheep in the millet subsystem can be explained by the adaptation of the ewes to the different partitioning of resources as they grow, reach maturity and then senesce. Overall, however, the effects of parity on parturition interval are slight in sheep in the millet subsystem and do not affect sheep in the rice subsystem. The parturition intervals for goats are not affected by parity in either subsystem. The significant effect of parity on annual reproductive rate is confined to goats in the millet subsystem, resulting from increased litter size with advancing age of the dam.
The effects of year are difficult to interpret. It is possible, however, that the maximum annual reproductive rate in goats in the millet subsystem during the middle period of the current study is related to the recovery of animals and food resources from the effects of the 1968-1973 Sahel drought, followed by a decline due to the effects of lowered rainfall during the early 1980s and increased pressure on feed resources as a result of a build-up of stock numbers. Why the same phenomenon is not significant in sheep in the same subsystem is not clear.
Although the effects of flock are significant for most variables, it has not yet been possible to isolate the specific mechanisms which cause the differences. In traditional systems in Africa, access to the basic resources of grazing and water is theoretically equal for all flocks. It is possible that individual management abilities play a role in acquiring better use of these resources by ensuring that animals have adequate time at grazing and are given sufficient water on a regular basis. Other practices such as keeping night-holding pens or picket areas free of manure, and simple veterinary interventions, may also influence flock reproductive performance.
A comparison of these results with those available from similar studies indicates that traditionally managed goats and sheep in Mali have a reproductive performance close to the African average. Observed litter sizes for goats range from 1.23 in Kenya (Wilson et al, 1984) to 1.51 in Sudan (Wilson, 1976c) and for sheep from 1.05 in Kenya to 1.14 in Sudan (Wilson and Clarke, 1976b). In Sudan, goats have been recorded as having parturition intervals of 238 days and in Kenya of 306 days. Intervals for sheep range from 275 days in Sudan through 301 days in Niger (Haumesser and Gerbaldi, 1980) to 320 days in Kenya. In other studies where goats and sheep have been recorded together under the same conditions, goats have always shown clearly superior annual reproductive rates. In central Mali, more detailed research, particularly on management skills, is needed to explain the superiority of sheep over goats.
In the systems studied there appears to be little possibility of improving total reproductive performance by attempting to shorten the parturition interval. Evidence from other tropical areas indicates that the figures recorded in this study are optimal. Creole goats in Guadeloupe average 237 days and the Indian Malabar 300 days (Garcia and Gall, 1981). Blackbelly sheep in the West Indies average 248 days and Pelibuey-West Africa sheep in Central America have an average interval of 245 days (Fitzhugh and Bradford, 1983).
Table 39. Data from the polynomial analyses of reproductive characteristics of goats and sheep in central Mali.
|
Species and trait |
System
|
Variable
|
Orthogonal regression |
Regression constants |
||||
|
a |
b |
c |
d |
|||||
|
Sheep |
||||||||
|
|
Litter size |
Millet |
Season |
Linear |
1.028 |
-0.0191 |
- |
- |
|
|
Parturition interval |
Millet |
Season |
Quadratic |
249.0 |
2.458 |
14.65 |
- |
|
|
|
|
Year |
Cubic |
277.2 |
5.711 |
-4.93 |
-4.91 |
|
|
|
|
Parity |
Quadratic |
256.8 |
3.077 |
8.38 |
|
|
|
Annual reproductive rate |
Millet |
Season |
Quadratic |
1.64 |
-0.0169 |
-0.0978 |
- |
|
Goats |
||||||||
|
|
Litter size |
Millet |
Season |
Linear |
1.151 |
-0.501 |
- |
- |
|
|
|
Parity |
Linear |
1.151 |
0.0102 |
- |
- |
|
|
|
Rice |
Year |
Cubic |
1.172 |
0.0686 |
-0.0185 |
-0.0347 |
|
|
|
|
Parity |
Linear |
1.135 |
0.1180 |
|
|
|
|
|
Parturition interval |
Millet |
Year |
Quadratic |
270.6 |
-22.73 |
6.764 |
- |
|
|
Annual reproductive rate |
Millet |
Year |
Quadratic |
1.700 |
-0.0231 |
-0.0682 |
- |
|
|
|
Parity |
Linear |
1.560 |
0.1392 |
- |
- |
|
Best-fit estimates from the polynomial analyses of reproductive characteristics of sheep and goats in central Mali.
Parturition interval (millet subsystem)
Annual reproductive rate (millet subsystem)
1) MS = millet subsystem; RS = rice subsystem.
Figure 44. Total number of parturitions per month and average number of young per parturition per month for goats and sheep in central Mali, 1978-1983.
The potential for improving the annual reproductive rate by increasing litter size seems more feasible. Where selection has been practised, the number of kids per birth for Creole goats in Guadeloupe was 2.33, for the Damascus in Israel 1.76 and for the Beetal and Jamnapari in India 1.70 and 1.45 respectively (Garcia and Gall, 1981). Sheep litter sizes of 1.84 in the Barbados Blackbelly and 1.24 in the Pelibuey in the Caribbean have been recorded (Fitzhugh and Bradford, 1983). The possibilities of increasing litter size by crossing with the more prolific African types from similar environmental areas might also be considered.
Manipulating flock structure so that as many of the more productive females of older parity as possible are maintained, could also improve reproductive performance. Encouraging first parturitions at as early an age as feasible and selecting breeding stock on the basis of performance at first parturition (Ozakuma et al, 1982) would also boost the overall annual reproductive rate.
The positive correlation of the number of parturitions with day length found in this study is contrary to that found in seasonally breeding sheep in temperate latitudes and is also contrary to the postulate of Hafez (1951; 1952) that in the tropics, the breeding season of sheep is not affected by light i.e. day length. The pattern of births in this study is, however, similar to that recorded in Chad where 60 to 80% of births occurred from November to February (Dumas, 1980). The principal climatic source of variance is ambient temperature, this phenomenon also having been noted previously (Lees, 1971). The combined, interacting effects of temperature and day length on oestrus activity of sheep have also been noted in Niger (Yenikoye et al, 1982). No previous studies appear to have attempted to separate the effects of a simple conception from those of litter size.
The lack of significant correlations with rainfall is rather surprising. In cattle in this area, parturition is very highly correlated (r = 0.56, d.f. = 70, P<0.001) with rainfall 9 months previously (Section 7). As rainfall is almost uniquely responsible for primary production - except for the appearance of some browse - and as almost no supplementary feeding is practiced in the systems studied, it does not appear that nutritional state is the proximal factor in the conception rate although it probably has more influence on the ovulation rate. The similar patterns of conception and of distribution of litter size in both goats and sheep are also puzzling in view of their different dietary habits.
More detailed experimental studies under controlled environmental conditions are required to explain fully the factors affecting fecundity and prolificacy in tropical goats and sheep.
Table 40. Number of parturitions per breeding female by age class.
|
Age by dentition class |
Number of parturitions |
|||||||||||||||||||
|
|
Sheep |
Goats |
||||||||||||||||||
|
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
0 |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
||
|
Milk teeth |
9 |
7 |
|
|
|
|
|
|
10 |
5 |
|
|
|
|
|
|
|
|
|
|
|
Permanent incisor pairs |
||||||||||||||||||||
|
|
1 |
8 |
29 |
2 |
3 |
|
|
|
|
42 |
62 |
5 |
2 |
|
|
|
|
|
|
|
|
|
2 |
|
10 |
4 |
5 |
3 |
|
|
|
4 |
35 |
19 |
3 |
|
|
|
|
|
|
|
|
|
3 |
1 |
3 |
6 |
6 |
4 |
1 |
|
|
2 |
22 |
39 |
12 |
2 |
1 |
|
|
|
|
|
|
|
4 |
|
|
9 |
8 |
13 |
4 |
2 |
1 |
2 |
5 |
33 |
52 |
40 |
24 |
15 |
4 |
1 |
2 |
2 |
|
All sheep |
18 |
49 |
21 |
22 |
20 |
5 |
2 |
1 |
60 |
129 |
95 |
69 |
42 |
25 |
15 |
4 |
1 |
2 |
2 |
|
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}