![]()
![]()
![]()
A.B. Charles
Department of Animal Production
University of Nairobi
P.O. Box 29053 Kabete Kenya
Introduction
Phenotype
Sex
Maternal environment
Climate
Season
Man
Compensatory growth
Conclusion
Summary
Phenotypic weight of mature females from a representative sample of African environments range from 30-45 kg for ewes and from 20-35 kg for does. Growth rates for the same sample range from 75-140 g/day for lambs and 50-100 g/day for kids, from birth to three months of age. This drops to between 30 and 75 g/day for both species at 3-12 months. Transformation of these to specific growth rates shows that much of the variation is due to differences in mature size. Sexual dimorphism is similar to that in temperate environments at birth, but less at maturity. Susceptibility to heat stress varies greatly within species and possibly between species. Seasonal differences have large and complex effects that often interact with other performance parameters. Differences between owners show that the human influence may be as important as any other factor. Compensatory growth may be affected by the stage of growth at which compensation is occurring.
Several major changes occur as an animal passes from the zygote to its mature form and size. Perhaps the most obvious change is in size and mass - these have been termed growth. In addition to these, there are fundamental changes in shape and body composition which have been termed differentiation. Because growth and differentiation are inseparable their combination is called development.
Growth and development are essentially ecological responses and it is thus appropriate to consider the factors affecting their outcome in an ecological framework. Firstly there is the genotype. Factors of environmental origin can be attributed to the internal (or physiological) and external environments. The former comprises principally the hormones affecting growth. The latter is usually subdivided into the physical, nutrient and biotic components from which man is usually omitted. Man's effects are so important and widespread that it is more convenient to consider him separately rather than to include him amongst the biotic components. In this paper attention will be confined to growth, for which most data are available, and in a form that permits a greater degree of comparison. Of the factors affecting growth, nutrition and disease are considered in other papers in these proceedings and so attention will be confined to the remaining factors.
The available data are inadequate for precise comparisons of the different genotypes of African sheep and goats and very few studies have been made where genetic effects have been separated from those of environmental origin. However this is probably less important at the present stage of development of Africa's small ruminant production systems than quantifying the phenotypic differences of different systems. Meagre though the data are, some broad comparisons are illuminating. Figure 1 is a comparative presentation of the mature weights of ewes and does for a selected sample of African breed types. The range of 30 to 45 kg covers the ewes, while does are rather smaller falling between 20 to 35 kg. No particular trend is suggested across the regions, or between the humid and arid zones.
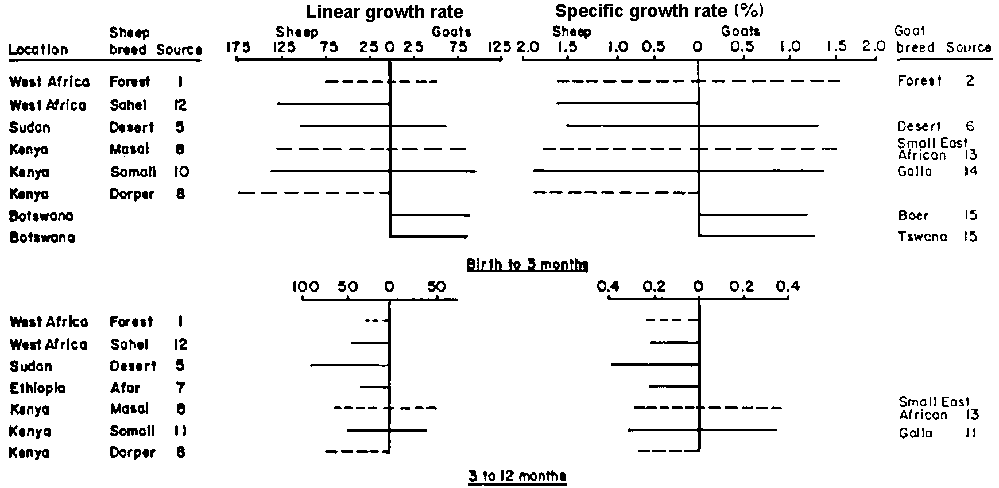
Figure 2 gives the gross and specific growth rates for periods between birth and 12 months for the same sample, together with two goat types from Botswana. Between birth and three months, the average daily growth rates (ADO) of sheep appear to range from 100 to 140 g, except for the two extremes - the very small West African Dwarf type and the improved Dorper. Goats have lower rates, with an ADG ranging from 50 to 100 g. Again no particular trend is apparent. When the comparisons are made on the basis of the specific growth rates, thus adjusting for differences in body size, differences are very much smaller amongst both sheep and goats, although goats are again lower ranging from 1.2 to 1.6, as compared with 1.5 to 1.9 for sheep.
Figure 1. Mature weights (kg) of female small ruminants from dry (____) and wet (----) areas
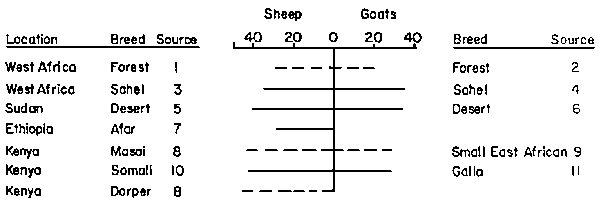
Sources:1. Akinkuolie, 1974
2. Matthewman, 1977
3. Wilson, 1983
4. Wilson, 1982
5. Wilson, 1976a
6. Wilson, 1976b
7. Wilson, 1980
8. Chemitei et al, 1975
9. Wilson, Peacock and Sayers, 1983
10. Rakoczi, 1974
11. Blackburn and Field, 1985
Sources1-11. as for Figure 1
12. Ferguson, 1964
13. de Haas and Chemitei, 1973
14. Chemitei, 1976
15. Ministry of Agriculture, Botswana, 1977* Note: The specific growth rate = ((logeW2 - logeW1)/(T2-T1)) x 100, where W1 and W2 are the weights at the ages T1 and T2
For the period from three to twelve months the ADG drops to 30 to 75 g for sheep and approximately 50 g for goats. The main exception is the Sudan Desert sheep which exhibits a higher ADG, suggesting either an element of compensatory growth for the group studied, or a slower rate of maturing.
Of course there is great variation. These levels can only be considered as examples under existing production systems, the performance of which can fluctuate widely. Certainly they are not an indication of genetic potentials: almost all present evidence indicates that these are far above existing levels. Where herds are small it is likely that sufficient inbreeding takes place to cause inbreeding depression. A more frequent exchange of sires among breeding groups could make a dramatic change unless an infectious breeding disease is present.
In a number of instances in the more humid zones exotic genotypes have been introduced, particularly with sheep. In the high altitudes of Kenya the Romney Marsh and Down breeds have shown that fat lamb of the same quality as in temperate areas can be produced from fifth and subsequent back crosses to these breeds (Macharia et al, 1975). The Dorset Horn has made a number of contributions to crossbreeding. Trail & Sacker (1969) report the results of one of these with the East African Blackhead. The growth of half-brads did not differ from indigenous lambs until post-weaning, when the growth rate was twice as great (both types suckled the indigenous ewe). A comparison of 75 per cent, 62.5 per cent and half-bred lambs showed no difference between the first two, but the half-brads were 26 per cent lighter at both weaning and 11 months of age. This indicated that the three-quarter and five-eighth breds were constrained by the environmental potential, which conforms with a number of findings that many tropical environments will not support much more than half-bred exotic crosses.
Perhaps the most extensive crossbreeding has been with the Dorper. It has been found that in a wide range of semi-arid tropical environments these cross-brads, and even the Dorper itself, have thrived, thus appreciably improving productivity (de Haas, Chemitei & Smith, 1973; Department of Agriculture, Zambia, 1973; Rudert, 1976). In the case of goats the Boer (Afrikander) has played a similar role (Kyeyune-Sendagi, 1970; de Haas & Chemitei, 1973; Department of Agriculture, Zambia, 1977; Mchau, 1979).
The degree of sexual dimorphism at birth, expressed as the male to female weight ratio, varies over the range 1.03-1.05 for sheep (Fitzhugh & Bradford, 1983), although Trail & Sacker (1969) reported a ratio as low as 1.02 for the East African Blackhead and Blackburn & Field (1985) report a ratio of 1.11 for Galla goats.
Data on mature weights of males are scarce. Those that are available generally indicate that ratios are in the range of 1.3-1.4 for both sheep and goats (Bradford, 1983; Charles, 1985), somewhat lower than the usual figure of 1.5 for temperate environments, although Bradford (1983) also reported a ratio of 1.6 for Sahel sheep in Senegal. The large variability with these mature ratios is probably due to varying levels of stress arising principally from the effects of uncontrolled breeding. The lower ratios indicate that males suffer more than females. This is supported by Charles' (1985) observations that the heaviest mature animals were castrates and the mature body weight ratio for castrates/females was approximately 2.0.
Aseasonal breeding imposes a heavy stress on females whose lactation peaks occur in the dry season and to a lesser extent upon those in late pregnancy at this time.
The major component of the maternal environment is the milk yield but mothering ability includes a number of others, probably of greater significance for the physical environment of the young. Unfortunately insufficient examination has been made of mothering ability in African sheep and goat systems to throw any light on these other aspects.
A practice that is fairly common among the pastoralists of eastern Africa does warrant attention. This is the separation of the new born young from the dam and maintaining it in a small basket-like construction which is often raised 1-2 m from the ground, until some six weeks of age. The young is allowed to suckle before going out to pasture in the morning and on return in the evening but otherwise remains in the basket. The technique is designed to improve survival and indeed is so effective during the first two months that mortality is the lowest for any age group, in contrast to the situation when the young run with their dams from birth in more favourable environments. However a considerable price is paid in lower growth rates. In Rendille these are so depressed that at six weeks of age the young have only doubled their birth weight (Caries, 1985). This is almost certainly due to the low level of milk intake but it is likely that other components also contribute.
A major component of the physical environment is the climate. Its effects upon growth are brought about by a complex of interacting components resulting in both direct and indirect effects upon the growing animal. The indirect effects are usually the more important for the ruminant: they include ambient temperature and factors controlling the level of soil moisture available for plant growth thus affecting the quantity and quality of available nutrients. Much the same complex of factors determines the microclimate of many micro-organisms and their vectors, these playing a major role in the dynamics of parasitic organisms and hence the levels of disease.
Direct effects upon small ruminants determine the extent to which any stress occurs due to extremes of temperature. In the tropics heat-stress predominates, resulting in a chain of events that depresses growth. Unfortunately detailed studies of heat-stressed African sheep and goats are very few. Symington (1960) carried out some comparative studies on Merino, Blackhead Persian, and the Long-fat-tailed sheep of Zambia. Under his conditions the Merino was the most heat adapted, the fat-tailed sheep being susceptible to the levels of heat stress imposed. Undoubtedly there is considerable variation among the many types of sheep and goats across Africa. There is, for example, an apparent difference between Small East African kids and Somali lambs in Rendille, in shade seeking at midday, with the latter demonstrating a strong shade preference not shown by the former.
Cold, wet and windy conditions, most serious in the high altitude areas, cause acute stress leading to hypothermia of the neonate, or pneumonia in suckling young and immatures. It is usually not sufficiently persistent to affect growth in the long term.
A number of workers have examined the short term effects of seasonal changes upon growth. Most have examined the seasonal effects around the time of birth and almost all have found that these have had major influences upon growth. Trail & Sacker (1966) observed that the occurrence of a dry season during stages between late pregnancy and weaning at five months had the worst effect during the early stage of suckling (from birth to two months) although when they examined the effects of month of birth with the East African Blackhead x Dorset Horn these were significant but too complex to permit a straight forward explanation.
In Zimbabwe Blackhead Persian and Dorset Horn cross-brads exhibited the beneficial effects of early grass growth coinciding with lactation but this could also lead to serious damage due to grass awns at a later stage if growth continued during the seeding period (le Roux, 1970).
Wilson, Peacock & Sayers (1983) showed that the four main seasons in Kenya Masailand (short rains, short dry, long rains, long dry) produced large differences in growth rates. For both sheep and goats the optimal time for growth was birth in the short dry season preceding the long rains.
Blackburn & Field (1985), from observations in a very arid part of Kenya (Rendille) also concluded that lambs grew faster when born one to two months before the rains, but this was not so apparent with kids. Working in the same area with goats, Carles (1985) concluded that birth just before or just after the rains was most favourable. Blackburn (1984) using simulation analysis examined a time period of the Rendille sheep system comprising two normal years separated by a drought year and concluded for such a period that aseasonal births would be preferable in terms of flock performance.
Season is a composite of a number of environmental effects: in the tropics the most obvious is the relation of rainfall with pasture growth. However the effects upon the incidence of infectious disease and temperature stress must not be overlooked. In effect the relieving of nutritional stress with the onset of the rains, often increases the stress from these other sources, so that the final outcome may not be easy to predict. There are additional correlated effects at subsequent stages of growth (e.g. the beneficial effect of early pasture growth for the young lamb may be correlated with the detrimental effect of grass awns post-weaning).
There is certainly sufficient variation in the response to seasonal differences to necessitate each main environmental type being considered on its own. Progress in predicting seasonal effects is going to depend upon increasing the precision with which the variable chosen for describing seasonal change does so.
It should also be noted that growth must not be considered in isolation and the final interpretation must also be based upon the levels of fertility and mortality as well as the body weights of the breeding females. When this was done by Wilson, Peacock & Sayers (1985) they concluded that with sheep the optimal combination remained the same, but the worst combination changed. In the case of goats the differences narrowed to the extent they were no longer significant.
In addition to short term effects there are evolutionary changes that have occurred in response to the extreme variation in food supplies across the seasons of the year. The most general adaptations to this situation have been reduced growth rates, a reduction in mature weight and the ability to lay down large fat reserves. Mature weight may be independent of height, as long limbs have often been developed to increase mobility, which is usually advantageous in such environments. The subcutaneous fat deposit, when confined to the tail and rump (and to some extent in the neck region) may be a response to heat stress.
The significance in the present context is that while these adaptations are eminently suitable for survival they conflict with the patterns of development required for meat production. As the latter is a lower priority, the environmental constraints must be relieved before much improvement in meat production is possible.
The human factor has only rarely been included, amongst the others considered so far, in the examination of sources of variation of growth. For domestic animals this is strange oversight. Fortunately this position is now changing and investigators are more frequently including man and his sociology amongst the factors they consider in their programmes for monitoring productivity of livestock. Wilson, Peacock & Sayers (1985) found large differences in au performance traits that were monitored among different herds. The ratio between the best and worst herds for a composite production index was 1.63 and 1.49 for goats and sheep respectively. They concluded that for most traits management was the factor responsible for most variation in performance.
Generally in the past sheep and goats have been a neglected and quite undeveloped resource in Africa. However the pressures upon resources that are developing today are changing this, and much more attention is being paid to them. This also behaves us to pay far more attention to the hum-an factor.
The phenomenon of compensatory growth has been known for long, whereby the depression of growth due to some stress may be compensated in whole or in part when the stress is removed. This ability plays a major role in modifying the various stressful effects on growth as imposed by the environment.
In most tropical ecosystems it appears that levels of stress are sufficient to produce permanent stunting as well as slower growth rates. In addition to this, evidence obtained by Carles (1985) suggests that there may be some interaction between the stage of growth experiencing stress and the subsequent pattern of compensatory growth. He observed with goats in Rendille that improved nutritional conditions at 7-9 months of age increased growth rates, but if the improvement was at 4-6 months of age there was an interaction with helminth burdens. If helminth burdens were negligible then the improvement in growth rate was greatest but mature size was decreased, relative to animals that did not experience either of these benefits. If the helminth burden remained then growth rates were increased less, growth was prolonged, and the mature size was appreciably greater. The fact that this interaction was associated with improved nutrition at 4-6 months and not at 7-9 months suggests that the earlier stage of growth may be affecting the outcome. In view of this one wonders to what extent the very severe stunting at the stage of growth immediately following birth may be responsible for the subsequent abnormal pattern of growth.
It is apparent that the growth of sheep and goats in the tropics is susceptible to as wide a range of influences as elsewhere. However there are many areas where the data are meagre and our understanding is extremely limited. Even knowledge of the current levels of performance of many of the tropical phenotypes is woefully inadequate. The effects of the complexes of season and the human factors and the patterns of compensatory growth all warrant particular attention.
Finally it should be noted that in terms of productivity of the systems, any changes in growth need to be interpreted in the light of correlated changes of the other main performance traits. This would be assisted by research into developing appropriate production indices that incorporate correct measures of all performance traits.
West African long-legged goat, adapted to increased mobility

![]()
![]()
![]()
{kind=link}