![]()
![]()
![]()
Abstract
Introduction
Principles of the method
Reference crops
References
S.K.A. Danso1, G. Hardarson2 and F. Zapata2
1 Joint FAD/IAEA Division, Wagramerstr. 5, POB 100, A-1400 Vienna, Austria
2 FAO/IAEA Agricultural Biotechnology Laboratory, A-2444 Seibersdorf Austria.
A great advantage with the use of 15N isotope dilution technique to estimate N2 fixed is its ability to give an integrated estimate of N fixation over a growing season or longer. It is the only method that can distinguish between soil, fertilizer and fixed N in field-grown crops. The technique has thus been extensively employed to quantify N-fixation in forage legumes, and there is an ever increasing interest in its use, as shown by recently published literature. Estimates made using the isotope dilution technique have established that most forage legumes derive a large proportion of their N from fixing atmospheric N2 and, in general, this is in excess of 70 - 80 % of their total N requirements.
The determination of the level of N-fixation by the 15N isotope dilution technique requires an assessment of the 15N/14N ratio in soil. This assessment is made by selecting an appropriate reference crop to assess the soil's 15N/14N ratio. Errors associated with using poor reference crops may be high at low values of fixation but become negligible at high values of fixation. With such a high proportion of the N in forage legumes generally coming from fixation therefore, the effect of a poor choice of a reference crop on N2 fixation estimate is low. Furthermore, the evaluation of the qualitative effect of treatments on N fixation requires no reference crop, if the treatments being compared received the same amount and enrichment of 15N-labelled fertilizer. They can be ranked simply by comparing the 15N enrichments within the legumes; the lower the 15N/14N ratio, the better the fixer it is. Only a quantitative determination of the magnitude of the effect requires use of a suitable reference crop.
The criteria that need to be observed in selecting suitable reference crops include: lack of N2-fixing ability, N-uptake patterns of the legume and the reference crop should match closely, similar periods for planting and harvesting, and proper sampling procedures.
In addition to forage legumes serving as rich sources of proteins for animals, there is currently a great interest in the use of legume-based forages in cropping systems for maintaining soil fertility and controlling erosion. Although many legumes have been shown to derive large amounts or proportions of their N from fixation (Walker et al, 1956; Henzell et al, 1968; Boddey et al, 1984: Heichel et al, 1981; Vallis et al, 1967; 1977, Heichel et al, 1985; West and Weding, 1985), it is also known that several factors can influence how much N2 is fixed in forage legumes (Vallis et al, 1967; Lynd et al, 1984; Carter and Sheaffer, 1983; Allos and Bartholomew, 1955; Chu and Robertson, 1974; Yamanaka and Holl, 1984). To exploit this natural and inexpensive source of N for optimum forage yields more effectively, therefore, reliable methods are needed to quantify biologically fixed N in forage legumes.
Various methods have been used to provide estimates of N2 fixed in forages, each of which has its own merits and disadvantages. Nodule number, weight and plant dry weight are among the earliest, most inexpensive and simplest methods (Bowren et al, 1969; Bell and Nutman, 1971; Heichel and Vance 1979; Heichel et al, 1984). Nodule parameters and plant yield, however, provide only indirect evidence of the extent of N2 fixation. They do not give a measure of how much N2 is fixed, neither do they always give an accurate indication of how much N2 is fixed (Danso, 1985).
The total N difference between a fixing and a non-fixing crop has been extensively used to estimate N2 fixed in various legumes (Williams et al, 1977; Boddoy et al, 1983; Broadbent et al, 1982; Phillips et al, 1983; Legg and Sloger, 1975; Bell and Nutman, 1971; LaRue and Patterson, 1981). The non-fixing crop is used to assess the amount or portion of the fixing crop's total N that came from soil. The assumption therefore is that the two crops, which may even belong to vastly different species, take up similar amounts of soil N. As pointed out by Rennie and Rennie (1983) and Danso (1985), the validity of this assumption is doubtful in most cases. The method also lacks precision. Moreover, where soil or fertilizer N levels are relatively high, N2 fixation may be low enough to be obscurred by sampling, analytical and other experimental errors (dose et al, 1982). However some N-difference estimates in good agreement with 15N-determined values for fixation have been reported (Ham, 1978; Broadbent et al, 1982; Phillips et al, 1983; Legg and Sloger, 1975).
The acetylene reduction technique of Hardy et al (1968) has been widely employed to estimate the nitrogen-fixing capacity of forage legumes (Goh et al, 1978; Heichel and Vance, 1979; Vance and Heichel 1981; Heichel et al 1981; Legg and Sloger, 1975; Lynd et al, 1984; Yamanaka and Holl, 1984; Heichel et al, 1984). Although simple, inexpensive and sensitive, this technique suffers from serious drawbacks, particularly since it is only a short-duration enzyme activity assay which does not take into account the existence of diurnal and seasonal variations in dinitrogen fixation (Ayanaba and Lawson, 1977; Vaughn and Jones, 1976) and thus makes extrapolation of such instantaneous assays to total nitrogen fixed over a growing season questionable. In addition, conventionally, a ratio of 3 is used to convert acetylene reduced to amount of N2 fixed, but this ratio has been found to vary widely among plants and in different environments (Bergersen, 1970; Goh et al, 1978; Hudd et al, 1980; Rennie et al, 1978; Knowles, 1981). Above all, the method is not well suited to field studies.
Since N2 is the substrate for N2 fixation, incorporation of 15N2 into plant tissues invariably provides the most direct and valid estimate of how much N2 is fixed. This technique, which was first used by Burris and Miller (1941), however, cannot be employed in field experiments, since it is not practicable to label all the N2 in the atmosphere in a normal field experiment (Fried et al, 1983). Its use has therefore been restricted to studies in closed chambers (Witty and Day, 1978; De-Polli et al, 1977).
However, by using the differences that exist in 15N enrichment of soil and the atmosphere (natural 15N-abundance technique), or by artificially inducing such a difference through the addition of 15N-enriched or depleted fertilizers to soil (in several reports called 15N-isotope-dilution method), it has been possible in the field to estimate directly the proportion or amount of N in a plant that is derived from symbiotic N2 fixation (McAuliffe et al, 1958; Vallis et al, 1967; Fried and Broeshart, 1975; Fried and Middelboe, 1977; Legg and Sloger, 1975; Kohl and Shearer, 1981; Bergersen and Turner, 1983; Broadbent et al, 1982; Heichel et al, 1985). According to Chalk (1985), more than 50 research papers on N2 fixation involving the addition of 15N enriched and depleted materials to soil have been published since 1975. Much of this interest in the technique is attributable to its applicability in the field and its ability to give an integrated estimate of N2 fixed over one or several growing seasons. The lowered cost of 15N-labelled fertilizers and the availability of cheaper instruments for 15N/14N-ratio analysis have also undoubtedly made a significant contribution. This paper will therefore be devoted largely to the use of 15N-enriched soils in N2-fixation studies.
The 15N-isotope-dilution principle stipulates that changes in 15N enrichment result when two sources that differ in N isotopic composition are uniformly mixed. The extent of change that results will depend on the magnitude of the differences in the initial enrichments of the individual sources, as well as the relative amounts of each. The isotope dilution equation for two sources may be written simply as:
(1)
a = [x (a1) + y (a2)] / (x+y)
where:
x = quantity of N fertilizer of higher enrichment with 15N abundance of a.
y = quantity of N fertilizer of lower enrichment with 15N abundance of a and
a = final 15N abundance of the mixtures of x + y.
If y is of natural abundance, then:
(2)
a = x (a1) / (x + y)
Thus, by adding 15N-enriched fertilizers to soil, plants take up N from soil with higher 15N composition than the approximately 0.3663 atom % 15N present in the atmosphere. The extent to which this higher 15N/14N ratio is lowered in a fixing plant by the lower 15N/14N ratio from the atmosphere is then a reflection of the magnitude of fixation.
It is therefore important to establish as accurately as possible the resultant 15N/14N ratio in the soil into which material with higher 15N/14N ratio has been incorporated. This, however, cannot be established easily by chemical extraction since the various soil sinks which provide N to the plant may have different 15N/14N ratios. Also, depending on how and when the 15N is applied, the ratio may not remain constant but may decline during the growing season, especially shortly after incorporation (Fried et al, 1983; Witty, 1983; Vallis et al, 1967). A reference crop that does not fix N2 has thus routinely been used to establish the integrated ratio of 15N/14N in soil over the whole growing season (McAuliffe et al, 1958; Vallis et al, 1967; Fried and Middelboe, 1977; Fried et al, 1983). This is represented in Figure 1. The original equation for estimating the proportion of N in a legume derived from N2 fixation as presented by McAuliffe et al (1958) involved first estimating the proportion of N in the legume derived from soil (X) from the ratio EL/EGL, where EL and EGL are the enrichments of 15N in the legume and grass, respectively. The proportion of N from fixation is then given by 1 - x1. This is essentially similar to the equation given by Fried and Middelboe (1977) for calculation of the percentage of N2 fixed as: atom % 15N excess in legume
(3)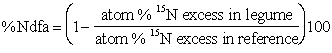
The use of natural 15N abundance for N2 fixation estimation requires that the 15N/14N ratio in the soil differs appreciably from that in the atmosphere, and is based on the same principles as above. It has been shown that many soils have higher 15N enrichments than the N2 in the surrounding atmosphere (Shearer et al, 1974; 1978; Cheng et al, 1964; Rennie et al, 1978; Virginia, 1980). Thus, soils seem to be naturally enriched in N. and the dilution of this by N2 derived from the atmosphere has been used to estimate N2 fixed in grain legumes (Amarger et al, 1979; Domenach et al, 1979; Ruschel et al, 1979; Kohl et al, 1980; Rennie, 1982; Rennie and Kemp, 1983) as well as forage legumes (Edmeades and Goh, 1979; Bergersen and Turner, 1983; Turner and Bergersen, 1983). A reference crop is again needed to establish the 15N/14N ratio in the soil. Given the usually small differences in 15N enrichments between soil and atmosphere, such differences are expressed in 0/00 units, i.e.,
Figure 1. Schematic diagram of the principle behind the use of the isotope dilution technique to estimate nitrogen fixation in crops.
(4)
d 15N = (R sample - R reference) / R reference
and
(5)
d 15N 0/00 = [(R sample - 1) / R reference] x 100
where
R = 15N/ (14N + 15N) and the reference is taken as the 15N enrichment in air, and is by definition equal to zero (Amarger et al, 1979).
Delwiche and Steyn (1970) have shown that isotopic fractionation of 15N and 14N occurs during biological dinitrogen fixation. In view of the high enrichments involved where 15N-labelled compounds have been added to soil, this isotopic discrimination between 15N and 14N normally does not affect the accuracy of the measurement or interpretation of results (Edmeades and Goh, 1979, Hauck and Bremner, 1976), while unless corrected for, significant errors could be introduced in the estimates of N2 fixed made by the natural abundance technique (Amarger et al, 1979; Mariotti et al, 1980; Kohl and Shearer, 1980). Isotopic fractionation can be assessed by growing legumes on sterilized sand watered with a nitrogen-free medium and estimating N isotopic composition in these plants dependent solely on N2 fixation. The new reference, 15NF is then given by:
(6)
d 15NF = d 15N air N + e
in which d 15N air is equal to zero and £ is the isotopic fractionation that occurred within the plant upon utilising atmospheric nitrogen. Nevertheless, because 15N/14N differences are small and discrimination values obtained experimentally vary from investigator to investigator, the method is still at a developmental stage.
In the 15N methods described above, the amount and enrichment of 15N material added to the legume and reference plots have to be the same. The amounts of N added often have to be small since high soil inorganic N levels normally inhibit N2 fixation (Butler and Ladd, 1985a; Allos and Bartholomew, 1955; Haystead and Marriott, 1979; Heichel et al, 1985). Under N-deficient conditions, however, the small amounts of N added so as to avoid interference in N2 fixation by the legume may be inadequate to support normal growth of the reference crop. By using the A-value approach of Fried and Broeshart (1975), it is possible to add different amounts and 15N enrichments to the legume and non-legume and still make valid estimates of N2 fixed. The basic concept behind this methodology is that plants will take up N from soil and any other sources in direct proportion to how much of each is available to the plants (Fried and Dean, 1952). This implies that the amount of N available in a given soil is the same for different crops exploiting N from a similar zone, and is also unchanged even under different fertilizer N levels (Aleksic et al, 1968). The A-value is estimated by the following equation of Fried and Dean (1952):
(7)
A = B (1 - y)/y
where A is the amount of a given nutrient available in, for example, soil (As), B is amount of fertilizer N added, and y is the proportion of N in the plant derived from fertilizer.
Thus for a crop like a legume which derives N from soil, fertilizer and fixation,
(8)
%Ndff / Af = %Ndfs / As = %Ndfa / Aa
where % Ndff, Ndfs and Ndfa refer to the proportions of N derived from fertilizer, soil and fixation, respectively.
The non-fixing crop estimates As, while the legume estimates Aa + As, and subtracting the latter from the former gives Aa. % Ndfa is then equal to
(%Ndff /Af) / Aa
where % Ndfa is the proportion of N derived from fixation.
Although the A-value approach has been a subject of much controversy (Broadbent, 1970; Boddey et al, 1983; Vose et al, 1982; Rennie and Rennie, 1983), several studies (Phillips and Bennett, 1978; Williams et al, 1977; Heichel et al, 1981; Wagner and Zapata, 1982) have reported good estimates made using this method. The A-value approach cannot, however, be used in mixed pasture stands unless a suitable external standard is used, as it involves the application of two separate rates.
Reports in the literature reveal that perennial ryegrass is about the most commonly used reference crop for estimating N2 fixation in forage legumes. Others that have been used include spear grass and Rhodes grass (Vallis et al, 1967), Pangola grass (Vallis et al, 1977), salt chess grass (Williams et al, 1977), barley (Witty, 1983), Dactylis glomerata (West and Wedin, 1985), tall fescue and red canary grass (Heichel et al, 1985).
Reference crops are needed solely for the assessment of the soil's
15N/14N ratio over the growing period. They are therefore
not essential when only qualitative differences between N2 fixed
in different treatments (which have received similar amounts 15N
enrichments of N fertilizer) are required. However, when accurate amounts or
proportions of N2 fixed are needed, a reference crop has to be used
and this constitutes the greatest potential source of error in the 15N
technique for determining N2 fixation in field experiments.
For example, with (NH)2SO4 as the source of 15N,
nodulated soya beans in Senegal were estimated to have fixed 71 kg N/ha in the
field relative to a non-nodulated soya bean control, while fixation by the same
plants with sudan grass as a control was estimated as 40 kg N/ha (Ganry, personnel
communication). The selection of an appropriate non-fixing crop is therefore
crucial under such circumstances, while, on the other hand, the error made by
using an improper reference crop may be negligible where the 15N/14N
ratio in soil remains constant during the period of growth, or where the proportion
of the plant's N that is fixed is very high, as is typical of many forage legumes.
In the case of established pastures, the non-fixing companion crop in the mixture,
if any, or the weeds growing together with the forage legumes is usually used
as reference crop. It is, however, desirable to establish their suitability
since these companion crops in some cases may not be appropriate as controls.
Fried et al (1983) list the following criteria which have to be considered in
assessing the suitability of a crop as a reference:
Absence of N2 fixing activity: Although at present
most forage legumes do not have suitable non-nodulating isolines, the absence
of substantial acetylene reduction activity in suitable crops is enough to ensure
that a potential reference crop does not fix N2 (Fried et al, 1983;
Phillips and Bennett, 1978).
Relative N uptake patterns: The N uptake profiles of the reference and
fixing-crop should match as closely as possible, since the 15N/14N
ratio of 15N-enriched soil does not normally remain constant but
usually declines with time (Vallis et al, 1967; Witty, 1983; Fried et al, 1983).
The basic assumption in using a reference crop therefore is that the two types
of crop will take up soil 15N/14N in the same proportion
throughout growth and, in the case of perennial forages, also within different
harvests, so as to reflect the changing soil 15N/14N ratio.
According to Witty (1983), even relatively small differences in N-uptake profiles
of fixing and non-fixing crops could lead to as much as 50% error when the rate
of change in soil enrichment is high. Unequivocal proof that the fixing and
non-fixing crops are deriving N of the same isotopic composition throughout
a growth cycle is, however, often difficult to obtain directly, and the few
N2-fixation studies that have attempted to establish this have done
so through indirect approaches (Witty, 1983; Fried et al, 1983; Wagner and Zapata,
1972). A time-course study of growth and nitrogen uptake is quite useful in
ascertaining that the potential reference crops do not deviate appreciably in
N uptake from the forage legumes of interest (Heichel et al, 1985). A suitable
solution to this problem is to label the soil in such a way that the 15N/14N
ratio does not change appreciably with time. In addition to the decline in 15N/14N
ratio with time, there is normally spatial variability of this ratio with soil
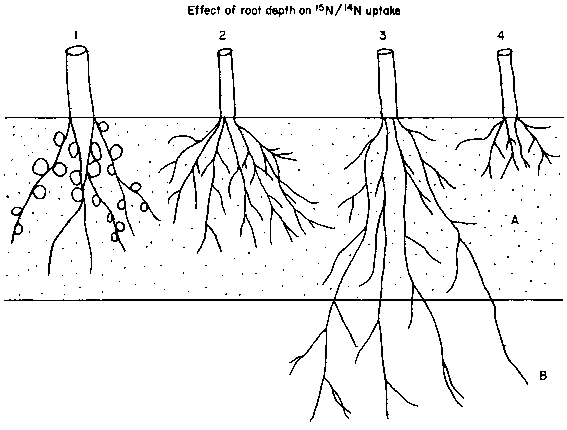
depth in field soils due to nonhomogeneous mixing (Ladd et al, 1981b). Thus,
although it has been pointed out by Fried et al (1983) that the essential prerequisite
is the absorption of N from a similar zone of incorporation (designated A in
Figure 2) as shown by crops 1, 2 and 4 in Figure 2, rather than from the same
depth, if the vertical gradient of 15N is substantial, problems could
arise by using crop 4, which could otherwise have been a good reference. Ideally,
the soil should be uniformly labelled to the depth that is explored by the plant
roots, as suggested by Boddey and Victoria (1985), but this is difficult to
achieve in a field experiment. Luckily, many forage legumes tend to be shallow
rooted, and it should not be too difficult to get equally shallow rooted non-fixing
plants to test for suitability as reference crops. In addition, a large proportion
of the available nitrogen is in the surface layer in most soils.
Figure 2. Diagramatic representation of the principles underlying the selection of an appropriate reference crop to assess soil 15N/14N ratio taken up by a fixing crop.
Relative effect of environment on reference and fixing crops:
In using reference crops to estimating N2 fixed in forage legumes,
it is essential to ensure that both fixing and non-fixing crops are adapted
to the climate of the area. Some forage legumes adapted to cool environments
may do poorly in warm environments and vice versa. Reference crops for cool-season
forage legumes should also be capable of growing under cool conditions, otherwise
the periods of growth/ establishment could be different and, as pointed out
by Fried et al (1983) and Witty (1983), while the adapted crop may grow early
and utilise much of the N of high 15N/14N ratio during
the initial period of N addition, the unadapted and late-developing crop will
only take up substantial amounts of N when the 15N/14N
ratio is quite low. An important criterion for the validity of the 15N
technique, that both fixing and non-fixing crop will take up N of the same isotopic
ratio throughout the period of growth, will then not be met if the soil's N
isotopic composition changes with time. For example, in an experiment on N2
fixation in Vicia faba during the cool season in Egypt, sudan grass,
wheat and barley were assessed for suitability as reference crops. Wheat and
barley, which are cool-season crops like Vicia faba, were found to be suitable
and gave a similar estimate of N2 fixed, while sudan grass, a warm-season
crop, was not suitable and gave a substantially lower figure for the amount
of N2 fixed (Hamissa, personal communication). This effect was, however,
negligible on the N2-fixation estimate made on faba bean, which derived
more than 80% of its N from fixation, but was large for chickpea, which fixed
less than 30% of its N. A late-developing reference crop could therefore underestimate
nitrogen fixation, and in low-fixing crops, negative values of estimated fixation
may be obtained (Witty and Ritz, 1984). In perennial crops, such as many forages,
the reference and fixing crops do not have to be adapted only to climatic factors
over one season, but possibly over several seasons and even years.
Time of growth of reference and fixing crops: In all N2 fixation
studies, it is essential that both the reference and the fixing crop are planted
and harvested together. Since many N2-fixation studies in forage
legumes have involved mixed crops, often both crops have been harvested together
even though the different crops have been separated from each other during or
after harvest. A few cases involving different planting dates due to, e.g. replanting
of one crop (Vallis et al, 1967) have been reported, and this should be avoided
as much as possible, since under such circumstances both the fixing and the
non-fixing crops may not be taking up N of the same isotopic composition throughout
growth (Fried et al, 1983).
Cropping systems: Forages may be grown alone (Simpson, 1965; Williams
et al, 1977; Butler and Ladd, 1985a; Phillips et al, 1983) or, as is the common
case, in mixtures with grasses (Vallis et al, 1967; Henzell et al, 1968; Broadbent
et al, 1982; Butler and Ladd, 1985b; Haystead and Lowe, 1977; Heichel et al,
1985). With sole forage legume cultivation, identical plots of a sole crop of
the reference species have often been established. In the case of stands of
legumes and non-legumes, an additional or optional choice is in using the non-fixing
crop within the mixture. However, competition by a non-legume growing in association
with forage legumes has been found to affect the growth and nitrogen uptake
of the latter (Vallis et al, 1967; Henzell et al, 1968; Butler and Ladd, 1985b).
Thus, in order for the use of a sole non-fixing crop to estimate N2
fixed in mixtures to be valid, it is essential to establish that the sole reference
crop and forage legume in the mixture (and not alone) have similar N uptake
and/or growth patterns. A second assumption is that the reference crop growing
alone and the forage legume in the mixture are sampling soil of similar 15N/14N
ratios.
However, since forage legumes often add more litter to the soil than do grasses
(Birch and Dougall, 1967; Vallis et al, 1977), decomposition of the forage legume
residues with lower 15N/14N ratios may lead to decreased
15N/14N ratios in soils under the mixed sward than under
pure grass stands (Heichel et al, 1984). Should this happen, it is not correct
to use a sole reference crop to estimate N2 fixed by a forage legume,
as they will be sampling soils of different 15N/14N ratios.
In adopting the companion grass or non-fixing crop in a mixed pasture to estimate
N2 fixed by the forage legume it is assumed that no significant N
transfer occurs from the legume to the non-fixing crop and that both the crops
are sampling similar 15N/14N ratios over time.
Many reports have shown that forage legumes transfer N to associated non-legumes
(Broadbent et al 1982; Bhaskar et al, 1984; Simpson, 1965). Broadbent et al
(1982) therefore concluded that isotope dilution methods are not suitable for
estimating N2 fixed in grass-legume mixtures. However, other reports
have shown that such N transfer does not occur or is not of significance (Vallis
et al, 1967; Butler and Ladd, 1985a; Haystead and Marriott, 1979; Haystead and
Lowe, 1977). Much of the evidence for N transfer has been obtained as a result
of the lowered 15N/14N ratio in a non-fixing crop grown
in association with a forage legume compared to the non-fixing crop grown alone
(Fried et al, 1983), with the lowered 15N composition being attributed
to the transfer of fixed N of lower 15N/14N ratio to the
associated grass. However, what has not been established in most of these studies
is whether the non-fixing crops under the two cropping systems had identical
growth (or N- uptake) patterns and were growing on soils with similar 15N/14N
ratios (Vallis et al, 1967). Evidence in the literature suggests that they may
have been growing on soils of different 15N/14N ratios
as a result of, e.g. differential litter accumulation (Birch and Dougall, 1967;
Vallis et al, 1977) or differences in mineralisation of native organic matter
under the two crops (Birch and Dougall, 1967; Vallis et al, 1977; Broadbent
and Nakashima, 1974), and the growth vigour (and thus possible U-uptake pattern)
of the non-fixing crop may have been altered as a result of growing near a legume
(Simpson, 1965; Vallis et al, 1967; Butler and Ladd, 1985b). These may help
to explain why in some studies the 15N enrichment in the non-fixing
crop grown alone was lower instead of higher than that of the non-fixing crop
in the mixture. Furthermore, lower 15N enrichments in mixed legume-grass
mixtures could be due simply to contamination of the separated grass roots-by
small amounts of legume root fragments (Butler and Ladd, 1985a).
As to the probability that non-fixing and fixing crops growing in association
may be sampling soil of different 15N/14N ratios, the
chances are not as high as when the reference and fixing crops are grown on
separate plots. This is because although higher litter decomposition due to
higher legume-litter turnover may result in a lowered 15N/14N
ratio in soil under mixed pasture than under grass alone, the altered soil 15N/14N
ratio, if homogeneous, is being sampled by both crops.
Sampling of plant material: It has been demonstrated that different plant
parts (e.g. seed, herbage, crowns and roots) of crops grown on 15N-enriched
soil may contain N of different isotopic composition from each other (Ladd et
al, 1981b; Rennie et al, 1978; Fried et al, 1983; Butler and Ladd, 1985a). This
difference in enrichment has been cited as a problem in the 15N-
isotope technique for estimating N2 fixed (Heichel et al, 1981; 1984).
However, as pointed out by Fried et al, (1983), errors due to differences in
15N enrichment can be minimised by sampling each of these plant parts
with fairly uniform N-isotopic composition separately for 15N determination
and using the following equation to derive a weighted atom % 15N
for the whole plant:
(10)
WPAE = [AE (a) x TN (a)+ AE (b) x TN (b)
+ AE (c) x TN (c) ] / TN (a+b+c)
Where
WPAE = weighted atom % 15N excess
AE = atom % 15N excess in different plant parts represented as a,
b and c.
TN = total N in parts a, b and c
In small-seeded forage legumes, it is even uncertain if differences in densities
of the small seeds and the rest of the plant are substantial enough to introduce
significant errors in sampling the whole plant. Perhaps these two approaches
need to be tested, since the whole plant approach avoids adding up errors from
each of the different samples and demands less labour and cost for separation
of plant parts and 15N/14N ratio analysis. Furthermore,
although the evidence for the existence of differences in 15N/14N
ratio in different plant parts is strong for N2-fixing legumes, results
obtained from the FAD/IAEA Biotechnology Laboratory in Seibersdorf, Austria
(Zapata et al, unpublished data) show that, while 15N distribution
in pods and straw were dissimilar for either Vicia faba or nodulated
soya bean, for non-nodulating soya bean and barley these parts were of similar
N-isotopic enrichment. Also, Haystead and Lowe (1977) and Heichel et al (1981)
have reported fairly uniform 15N enrichments in shoots, roots and
stubble of white clover and alfalfa, respectively. As a compromise, therefore,
it may be advisable to perform preliminary N-isotopic analysis on different
plant parts of crops used in an experiment and if the N-isotopic composition
is fairly uniform, there will then be no need to continue splitting the different
plant parts in the experiment as well as in subsequent ones. However, the problem
may not arise in many forage legumes since, in most cases, these are harvested
before seed production.
Most estimates of nitrogen fixation have been calculated on the basis of either
herbage or above-ground plant parts (Williams et al, 1977; Phillips and Bennett,
1978; Heichel et al, 1984). In many grain legumes, roots or underground structures
may not constitute a significant proportion of total dry matter, and therefore
the error made by excluding roots may not be as serious as in forage legumes,
some of which could have a large proportion of the total dry matter below ground
(Heichel et al, 1981; 1984). In such cases, not only would the estimate of the
total amount of N2 fixed be inaccurate, but the proportion of N2
fixed may be overestimated, since roots are known to have higher 15N
enrichments than foliage or seeds (Bergersen and Turner, 1983). In addition,
for many perennial legumes the N stored in these underground structures and
unharvested portions of foliage may, after decomposition exert a significant
effect on the dilution of the soil 15N/14N ratio. Errors
in N2 fixation estimates may thus result unless both the legume and
its reference crop have similar amounts of carryover N. which is not likely
under many circumstances. It is therefore advisable to estimate this stored
N and make a correction for its effect on the isotopic composition of N at harvest,
as suggested by Haystead and Lowe (1977), who proposed the following equation
for correcting for the effect of carryover plant N.
(11)
D N = Ep/(1-NR+O/Np)
where D N is the nitrogen uptake during the growing
period, Ep is the measured 15N enrichment of whole plants, Np
is the total nitrogen content of the whole plant at harvest and NR+O
is the nitrogen in stubble, roots and nodules. In harvesting roots, special
efforts have to be made to dig up as much of the roots as possible, since this
could be a source of error, and since any soil N adhering to the roots could
drastically alter the 15N/14N ratio in them thorough washing
is recommended.
Forms and methods of incorporation of 15N-labelled materials:
Many different formulations of 15N-labelled materials have been
used to estimate N2 fixation in grain and forage legumes. These include
(NH4)2SO4 (McAuliffe et al, 1958; Vallis et
al, 1967; 1977; Haystead and Lowe, 1977; Phillips and Bennett, 1978), NH4Cl
(Williams et al, 1977; Witty, 1983), urea (Hami 1978; Edmeades and Goh, 1979;
Steele, 1983), residual 15N in soil (Fried et al, 1983), organic
residues (Fried et al, 1983; Boddey et al, 1984; Witty and Ritz, 1984; Butler
and Ladd, 1985a), oxamide (Witty and Ritz, 1984; Hauck, personal communication),
as well as many others.
From the literature it seems that (NH4)2SO4
so has been most commonly used, although no explanation has been given for this
preference. However, Vallis et al (1967) observed a greater uptake of N from
K15NO3 than from (15NH4)2SO4
and suggested that ammonium would probably be better than the nitrate form of
fertilizer when the aim is to have much of the tracer incorporated in the internal
soil nitrogen cycle. On the other hand, nitrate is better when the objective
is to minimise microbial transformation before uptake. According to Haystead
and Lowe (1977), in order not to disturb the soil N cycle, labelled N should
be added in a form already present in the soil. Much more work on comparing
different 15N fertilizers and materials for measuring N2
fixation in different legume is needed, such as the rates of N release by different
formulations and their influence on the declining soil 15N/14N
ratio (Witty and Ritz, 1984), preferential uptake of different ionic forms of
N by different crops and how they affect N2-fixation estimates (Ledgard
et al, 1985b). Slow-release formulations and 15N-labelled organic
matter offer great promise in N2-fixation studies, since the 15N/14N
ratio in the soil would remain more stable due to the rather slow release of
a small but constant amount of N. which would probably lead to less errors between
different reference crops and legumes attributable to differences in relative
N-uptake profiles. This is supported by the finding of Butler and Ladd (1985a)
that the inorganic N released within a 12-week period from each of 4 soils into
which 15N-labelled organic matter had 15 been incorporated had approximately
constant 15N atom % enrichment. Similar results have also been reported
by Legg and Sloger (1975) and Ladd et al (1981b). In such instances the isotopic
enrichment of soil N taken up by legumes and non-fixing crops would be similar,
irrespective of any mismatch in growth patterns or 15N-uptake profiles.
Furthermore, Butler and Ladd (1985a) observed that the proportion of 15N
in the inorganic N released from 15N-enriched materials was high
enough to disregard isotopic discrimination effects during N uptake.
On the other hand, although Boddey et al (1984) did not get a constant 15N
enrichment of soil N until the fourth harvest in soil enriched with 15N-labelled
organic material, not only was the coefficient of variation of the first three
harvests lower for this treatment, but also there was a closer agreement between
the 15N-uptake profiles of the nodulated and non-nodulated plants
than when labelled inorganic fertilizer was applied.
Just as with the different forms, many methods of 15N application
have been used, including broadcast at planting (Ham, 1978), broadcast and incorporated
1 week before planting (Heichel et al 1981), sprayed or injected into soil as
solution (Simpson, 1965; Vallis et al, 1967; Fried et al, 1983; West and Wedin,
1985; Broadbent et al, 1982) and banding (Fried et al, 1983). The greatest problems
with the different methods of application may, however, be related to uniformity
of fertilizer distribution in the soil and how each method affects the rate
of release of 15N (or tie-up in the soil colloid or organic matter).
Certainly, surface application, unless followed by irrigation or rainfall, may
lead to vertical gradients in the 15N composition of soil which,
if substantial, may make it crucial to have roots of reference and fixing crop
at similar depth. Solution application has the advantage that small amounts
of 15N-labelled fertilizer (which would be difficult to distribute
evenly on the soil surface) can be dissolved in a large volume of water and
applied uniformly to cut down on spatial variability. Also, this approach makes
it possible to mix fertilizers of different 15N enrichments to obtain
a homogeneous N- isotopic composition.
Unlike N2-fixation studies in annual grain legumes, for which single
doses of 15N-enriched fertilizer have been applied usually at planting,
those involving forage legumes have often used equal-sized additions of small
doses of 15N fertilizer applied frequently, at short intervals (Catchpoole,
1983; Vallis et al, 1967; 1977; Haystead and Lowe, }977). This may have the
advantage that the 15N/14N ratio in the soil may be high
at most periods of the plant's growth and may thus help to reduce errors due
to the rapid decline in 15N/14N ratios that follows after
the application of 15N fertilizers (Fried et al, 1983; Heichel et
al, 1984; Chalk, 1985). The 15N enrichments of the inorganic fertilizers
used on forage legumes have ranged from as low as 3 atom % 15N excess
(Phillips et al, 1983) to as high as 97.6% (Haystead and Lowe, 1977). Excessively
high 15N enrichments increase the cost of the experiment unnecessarily.
An experiment was set up in the FAD/IAEA Agricultural Biotechnology Laboratory
in Seibersdorf, Austria, to test how different N rates, 15N enrichments
and frequency of fertilizer application affect estimates of N2 fixation
in alfalfa in consecutive harvests of an alfalfa-ryegrass mixture over a 2-year
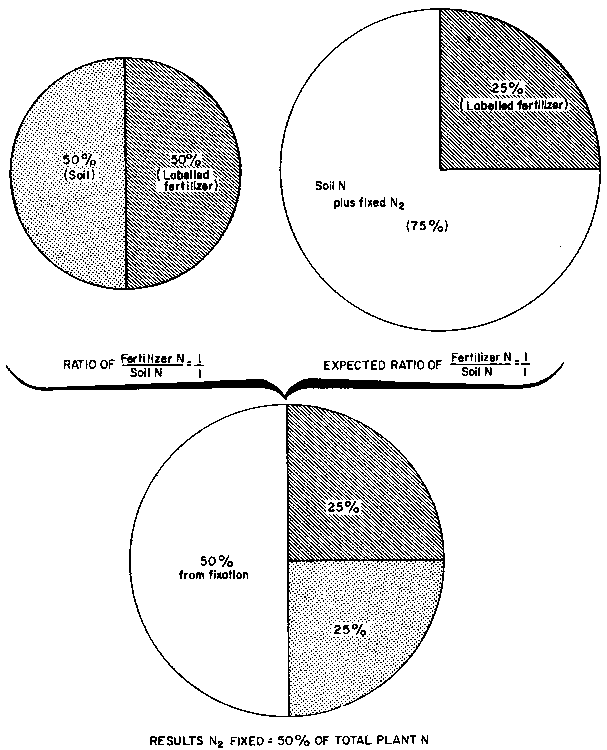
period. The treatments, which are represented schematically in Table 1, comprised:
1. Treatment 1, in which urea of high 15N enrichment (50 atom % 15N excess) was applied at a low rate (5 kg N/ha) at establishment and then to a new subplot after each harvest.Table 1. Effect of 15N enrichment, fertilizer level and method of application on N2 fixation estimates.
2. Treatment 2, where urea of low 15N enrichment (5 atom % 15N excess) was applied at a low rate (5 kg N/ha) to the same plot at planting and then after each harvest.
3. Treatment 3, which consisted of the application of a low rate of N fertilizer (5 kg N/ha) of a moderately high 15N enrichment (16.65 atom % 15N excess) once a year (instead of twice a year as in Treatments 1 and 2) to the same subplot.
4. Treatment 4, in which a high initial dose of urea of low 15N enrichment (45 kg N/ha, 5.73 atom % 15N excess) and none thereafter during the 2 years.
|
Treatment number |
Atom % Subplot |
15N excess |
kg N/ha |
|||
|
1st Year |
2nd Year |
|||||
|
T1 |
T2 |
T3 |
T4 |
|||
|
1 |
i |
50 |
5* |
5 |
5 |
5 |
|
ii |
50 |
5 |
5* |
5 |
5 |
|
|
iii |
50 |
5 |
5 |
5* |
5 |
|
|
iv |
50 |
5 |
5 |
5 |
5* |
|
|
2 |
i |
5.73 |
5* |
5* |
5* |
5* |
|
3 |
i |
16.65 |
5* |
5 |
5* |
5 |
|
4 |
i |
5.73 |
45* |
0 |
0 |
0 |
* = 15N-labelled fertilizer applied.
The results showed that all treatments gave similar estimates of N2
fixation, except Treatment 4, in which the high initial rate of N reduced N2
fixation (Hardarson, Danso and Zapata, unpublished). Since the cost of 15N-enriched
inorganic fertilizers rises disproportionately as the percentage of 15N
increases, the best treatment from this study would be Treatment 2 (application
of 5 kg N/ha of a 5 atom % 15N excess fertilizer (0.025 g 15N/m2)
at establishment and also after each harvest), followed by one in which 5 kg
N/ha of 16.65 atom % 15N excess urea is applied at establishment
and then after every other harvest.
Further trials by participants of the FAD/IAEA Coordinated Research Programme
(CRP) on the use of nuclear techniques in pasture management were carried out
(Table 2).
Table 2. Schematic diagram of effect of methods of 15N fertilizer
application on estimates of nitrogen fixation.
|
Treatment number |
Subplot |
Atom %15N excess |
Harvest |
||||
|
1 |
2 |
3 |
4 |
etc. |
|||
|
1 |
i |
5 |
5 |
5 |
5 |
5 |
|
|
ii |
5 |
5 |
5* |
5 |
5 |
|
|
|
iii |
5 |
5 |
5* |
5 |
5 |
|
|
|
iv |
5 |
5 |
5 |
5* |
5* |
|
|
|
2 |
i |
5 |
5* |
5* |
5* |
5* |
|
|
3 |
i |
10 |
5* |
5 |
5* |
5 |
|
* = 15N labelled fertilizer applied.
The preliminary results (unpublished) have confirmed that 5 kg N/ha of 5 atom
% 15N-enriched fertilizer is enough to measure N2 fixed
in forage legumes. The three different approaches were used (i) application
at planting and then to a new subplot after each harvest; (ii) application of
15N fertilizer at planting and then after each harvest to the same
subplot; (iii) application at planting, second harvest and then after every
second harvest to the same subplot. Each gave similar estimates of the proportion
of N derived from fixation. Most of the forage legumes studied fixed in excess
of 70 % of their total N requirements. Applying 15N fertilizer to
the same subplot has the advantage of confining the isotope experiment to a
small area. A major disadvantage, however, is that the isotopic-derived values
such as % Ndff and % fertilizer utilization cannot be reliably estimated after
the first harvest because of the unknown contribution from carryover and residual
15N in previous applications.
Aleksic Z. Broeshart H and Middelboe V. 1968. The effect of nitrogen fertilization on the release of soil nitrogen. Plant and soil 29: 474-478.
Allos H F and Bartholomew M V. 1955. Effect of available nitrogen on symbiotic fixation. Soil Sci. Soc. Am. Proc. 19: 182-184.
Amarger N. Mariotti A, Mariotti F. Durr J C, Bourguignon C and Lagacherie B. 1979. Estimate of symbiotically fixed nitrogen in field-grown soybeans using variations in 15N natural abundance. Plant and Soil 52: 269-280.
Ayanaba A and Lawson T L. 1977. Diurnal changes in acetylene reduction in field-grown cowpeas and soybeans. Soil Biol. Biochem. 9: 125-129.
Bell F and Nutman P S. 1971. Experiment on nitrogen fixation by nodulated lucerne. Plant and Soil, Special Volume. pp. 231-204.
Bergersen F G. 1970. The quantitative relationship between nitrogen fixation and the acetylene reduction assay. Aust. J. Biol. Sci. 23: 1015-1025.
Bergersen F. J. and Turner G L. 1983. An evaluation of 15N methods for estimating nitrogen fixation in a subterranean clover-perennial ryegrass sward. Aust. J. Agric. Res. 34: 391-401.
Bhaskar A, Vyas K G and Pandya S M. 1984. Nitrogen relationships of Vigna mango in pure stands and in Celosia argentea mixtures. Ann. Bot. 54: 799-805.
Birch H F and Dougall H W. 1967. Effect of a legume on soil nitrogen mineralisation and percentage nitrogen in grasses. Plant and Soil 27: 292-296.
Boddey R M and Victoria R L. 1985. Estimation of biological nitrogen fixation associated with Brachiaria and Paspalum grasses using 15N labelled orgainc matter and fertilizer. Plant and Soil: (in press).
Boddey R M, Chalk P M, Victoria R and Matsui K. 1983. The 15N-isotope dilution technique applied to the estimation of biological nitrogen fixation associated with Paspalum notatum cv. Batatais in the field. Soil Biol. Biochem. 15: 25-32.
Boddey R M, Chalk P M, Victoria R L and Matsui E. 1984. Nitrogen fixation by nodulated soybean under tropical field conditions estimated by the 15N isotope dilution techinque. Soil Biol. Biochem. 16: 583-588.
Bowren K E, Cooke D A and Downey R K. 1969. Yield of dry matter and nitrogen from tops and roots of sweet clover, alfalfa and red clover at five stages of growth. Can. J. Plant Sci. 49: 61-68.
Broadbent F E. 1970. Variables affecting A values as a measure of soil nitrogen availability. Soil Sci. 110: 19-23.
Broadbent F E and Nakashima T. 1974. Mineralization of carbon and nitrogen in soil amended with carbon-13 and nitrogen-15 labelled plant material. Proc. Soil Sci. Soc. Am. 38: 313-315.
Broadbent F E, Nakashima T and Chang G Y. 1982. Estimation of nitrogen fixation by isotope dilution in field and greenhouse experiments. Agron. J. 74: 625-628.
Broadbent F E, Rauschkolb R S. Lewis K A and Chang G Y. 1980. Spatial variability of nitrogen-15 and total nitrogen in some virgin and cultivated soils. Soil Sci. Soc. Am. J. 44: 524-527.
Burris R H and Miller C E. 1941. Application of 15N to the study of biological nitrogen fixation. Science 93: 114-115.
Butler J H A and Ladd J N. 1985a. Symbiotically fixed and soil-derived nitrogen in legumes grown in pots in soils with different amounts of available nitrate. Soil Biol. Biochem. 17: 47-55.
Butler J H A and Ladd J N. 1985b. Growth and nitrogen fixation by Medicago littoralis in pot experiments: Effect of plant density and competition from Lolium multiforum. Soil Biol. Biochem. 17:355-361.
Carter P R and Sheaffer C C. 1983. Alfalfa response to soil water deficits. III. Nodulation and N2 fixation. Agron. J. 23: 985-990.
Catchpoole Y R. 1983. Efficiency of legumes to supply nitrogen for soil/plant/animal system in mixed pastures with particular reference to the role of N. In: Nuclear techniques in improving pasture management. IAEA, Vienna, Austria. pp. 3 - 16.
Chalk P M. 1985. Review. Estimation of N2 fixation by isotope dilution: An appraisal of techniques involving 15N enrichment and their application. soil Biol. Biochem. 17: 389-410.
Cheng H H. Bremner J M and Edwards A P. 1964. Variations of nitrogen-15 abundance in soils. Science 146: 1574-1575.
Chu A C P and Robertson A G. 1974. The effects of shading and defoliation on nodulation and nitrogen fixation by white clover. Plant and Soil 41: 509-519.
Danso S K A. 1985. Methods of estimating biological nitrogen fixation. In: H Ssali and S O Keya, eds. Biological nitrogen fixation in Africa. Rhizobium MIRCEN, Nairobi, Kenya. pp. 224-244.
Delwiche C C and Steyn P L. 1970. Nitrogen isotope fractionation in soils and microbial reactions. Env. Sci. Technol. 4: 929-935.
DePolli H. Matsui E, Dobereiner J and Salati E. 1977. Confirmation of nitrogen fixation in two tropical grasses by 15N2 incorporation. Soil Biol. Biochem. 9: 119-123.
Domenach A M, Chalamet A and Pachiandi C. 1979. Estimation de la fixation d'azote par le soya a l'aide de deux methodes d'analyses isotopiques. C. R. Acad. Sci. Ser. D 289:291-293.
Edmeades D C and Goh K M. 1979. The use of the 15N-dilution techinque for field measurement of symbiotic nitrogen fixation. Commun. soil Sci. Plant Anal. 10: 513-520.
Fried M and Broeshart H. 1975. An independent measurement of the amount of nitrogen fixed by a legume crop. Plant and Soil 43: 707-711.
Fried M and Broeshart H. 1981. A further extension of the method for independently measuring the amount of nitrogen fixed by a legume crop. Plant and Soil 62: 331-336.
Fried M and Dean L A. 1952. A concept concerning the measurement of available soil nutrients. Soil Sci. 73: 263-271.
Fried M and Middelboe V. 1977. Measurement of amount of nitrogen fixed by a legume crop. Plant and Soil 47: 713-715.
Fried M, Danso S K A and Zapata F. 1983. The methodology of measurement of N2 fixation by non-legumes as inferred from field experiments with legumes. Can. J. Microbial. 29: 1053-1062.
Goh K M, Edmeades D C and Robinson B W. 1978. Field measurements of symbiotic nitrogen fixation in an established pasture using acetylene reduction and a 15N method. Soil Biol. Biochem. 10: 13-20. 15
Ham G E. 1978. Use of 15N in evaluating symbiotic N2 fixation of field-grown soybeans. In: Isotopes in biological dinitrogen fixation. IAEA, Vienna, STI/PUB/478. pp. 151-161.
Hardy R W F. Holsten R D, Jackson E K and Burns R C. 1968. The C2H2 -C2H4 assay for N2 fixation: Laboratory and field evaluation. Plant Physiol. 43: 1185-1207.
Hauck E D and Bremner J M. 1976. Use of tracers for soil and fertilizer nitrogen research. Adv. Agron. 28: 219-266.
Haystead A and Lowe A G. 1977. Nitrogen fixation by white clover in hill pasture. J. Brit. Grassl. Soc. 32: 5763.
Haystead A and Marriott C. 1979. Transfer of legume nitrogen to associated grass. Soil Biol. Biochem. 1: 99-104.
Heichel G H and Vance C P. 1979. Nitrate-N and Rhizobium strain roles in alfalfa seedling nodulation and growth. Crop Sci. 19: 512-518.
Heichel G H. Barnes D X and Vance C P. 1981. Nitrogen fixation of alfalfa in the seeding year. Crop Sci. 21: 330-335.
Heichel G H. Hardarson G. Barnes D K and Vance C P. 1984. Dinitrogen fixation, herbage yield, and rhizobial preference of selected alfalfa clones. Crop Sci. 24: 1093-1097.
Heichel G H. Vance C P. Barnes D K and Henjum K I. 1985. Dinitrogen fixation, and N and dry matter distribution during 4 year stands of birdsfoot trefoil and red clover. Crop Sci. 25: 101-105.
Henzell E F. Martin A E, Ross P J and Haydock R P. 1968. Isotopic studies on the uptake of nitrogen by pasture plants. IV. Uptake of nitrogen from labelled plant material by Rhodes grass and siratro. Aust. J. Agric. Res. 19: 65-77.
Hudd G A, Lloyd-Jones C P and Hill-Cottingham D G. 1980. Comparison of acetylene-reduction and nitrogen-15 techniques for the determination of nitrogen fixation by field beans (Vicia faba) nodules. Physiol. Planta. 48:111-115.
Knowles R. 1981. The measurement of nitrogen fixation. In: A H Gibson and W E Newton, eds. Current perspectives in nitrogen fixation. Australian Academy of Sciences, Canberra. pp. 327-333.
Kohl D H and Shearer G. 1980. Isotopic fractionation associated with symbiotic N2 fixation and uptake of NO3 -e by plants. Plant Physiol. 66: 51-56.
Kohl D H and Shearer G B. 1981. The use of soil lightly enriched in 15N to screen for N2 fixing activity in plants. Plant and Soil 60: 487-489.
Kohl D H. Shearer G and Harper J E. 1980. Estimates of nitrogen fixation based on differences in the natural abundance of 15N in nodulating and non-nodulating isolines of soybeans. Plant Physiol. 66: 61-65.
Ladd J N. Oades J M and Amato M. 1981a. Microbial biomass formed from 14C, 15N-labelled plant material decomposing in soils in the field. Soil Biol. Biochem. 13: 119126.
Ladd J N. Oades J M and Amato M. 1981b. Distribution and recovery of nitrogen from legume residues decomposing in soils sown to wheat in the field. Soil Biol. Biochem. 13: 251-256.
Ladd J N. Amato M, Jackson R B and Butler J H. 1983. Utilisation by wheat crops of nitrogen from legume residues decomposing in soils in the field. Soil Biol. Biochem. 15: 231-238.
LaRue T A and Patterson R. 1981. How much nitrogen do legumes fix? Adv. Agron. 34: 15-38.
Ledgard S F. Morton R. Freney J R. Bergersen F J and Simposon J R. 1985a. Assessment of the relative uptake of added and indigenous soil nitrogen by nodulated legumes and reference plants in the 15N dilution measurement of N2 fixation: Derivation of method. soil Biol. Biochem. 17: 317-321.
Ledgard S F. Simpson J R. Freney J R. Bergersen F J and Morton R. 1985b. Assessment of the relative uptake of added and indigenous soil nitrogen by nodulated legumes and reference plants in the 15N dilution measurement of N2 fixation: Glasshouse application of method. Soil. Biol. Biochem. 17: 323-328.
Legg J O and Sloger C. 1975. A tracer method for determining nitrogen fixation in field studies. In: E R Klein and P D Klein, eds. Proceedings of the 2nd international conference on_stable isotopes. US Energy Research and Development Administration, Washington. pp. 661-666.
Lynd J Q. Hanlon D A Jr and Odell G V Jr. 1984. Soil fertility and defoliation effects with arrowleaf clover and nitrogen fertilizer equivalence of crimson-arrowleaf clover combinations. Agron. J.- 76: 13-16.
Marriotti A, Pierre D, Vedy J C and Bruckert S. 1980. The abundance of natural nitrogen-15 in the organic matter of soils along an altitudinal gradient (Chabalais, Haute-Savoie). Catena 7: 293-300.
McAuliffe C, Chamblee D S. Uribe-Arango H and Woodhouse W W Jr. 1958. Influence of inorganic nitrogen on nitrogen fixation by legumes as revealed by 15N. Agron. J. 50: 334-337.
Phillips D A and Bennett J. 1978. Measuring symbiotic nitrogen fixation in rangeland plots of Trifolium subterraneum L. and Bromus mollis L. Agron. J. 70: 671674.
Phillips D A, Center D M and Jones M B. 1983. Nitrogen turnover and assimilation during re-growth in Trifolium subterraneum L. and Bromus mollis L. Plant Physiol. 71: 472-476.
Rennie R J. 1979. Comparison of 15N-aided methods for determining symbiotic dinitrogen fixation. Rev. Ecol. Biol. du 501. 16: 455-463.
Rennie R J. 1982. Quantifying dinitrogen fixation in soybean by 15N isotope dilution: The question of the non-fixing control plant. Can. J. Bot. 60: 856-861.
Rennie R J and Rennie D A. 1983. Techniques for quantifying N2 fixation in association with non-legumes under field and greenhouse conditions. Can. J. Microbiol. 29: 10221035.
Rennie R J and Kemp G A. 1583. Dinitrogen fixation in field beans quantified by 15N isotope dilution. I. Effect of strains of Rhizobium phaseoli. Agron. J. 75: 640-644.
Rennie R J. Rennie D A and Fried M. 1978. Concepts of 15N usage in dinitrogen fixation studies. In: Isotopes in biological dinitrogen fixation. IAEA, Vienna, Austria. STI/PUB/478 pp. 107-130.
Ruschel A P. Vose P B. Victoria R L and Salati E. 1979. Comparison of isotope techniques on nodulating and non-nodulating isolines to study the effect of ammonium fertilization on dinitrogen fixation in soybean, Glycine max (L). Merrill. Plant and Soil 53: 513-525.
Shearer G B. Kohl D H and Commoner B. 1974. The precision of determination of nitrogen-15 in soils, fertilizers, and shelf chemicals. Soil Science 118: 308-316.
Shearer G. Kohl D H and Chien S H. 1978. The nitrogen-15 abundance in a wide variety of soils. Soil Sci. Soc. Am. J. 42: 899-902.
Simpson R J. 1965. The transference of nitrogen from pasture legumes to an associated grass under several systems of management in pot culture. Aust. J. Agric. Res. 16: 915-926.
Steele K W. 1983. Quantitative measurements of nitrogen turnover in pasture systems with particular reference to the role of N. In: Nuclear techniques in improving pasture management. IAEA, Vienna, Austria. pp. 17-35.
Turner G L and Bergersen F J. 1983. Natural abundance of 15N in root nodules of soybean, lupin, subterranean clover and lucerne. Soil Biol. Biochem. 15: 525-530.
Vallis I, Haydock K P. Ross P J and Henzell E F. 1967. Isotopic studies on the uptake of nitrogen by pasture plants. III. The uptake of small additions of 15N-labelled fertilizer by Rhodes grass and Townsville lucerne. Aust. J. Agric. Res. 18: 865-877.
Vallis I, Henzell E F and Evans T R. 1977. Uptake of soil nitrogen by legumes in mixed awards. Aust. J. Agric. Res. 28: 413-425.
Vance C P and Heichel G H. 1981. Nitrate assimilation during vegetative re-growth of alfalfa. Plant Physiol. 68: 1052-1056.
Vaughn C E and Jones M B. 1976. Nitrogen fixation by intact annual rangeland species in soil. Agron. J. 68: 561564.
Vose P B. Ruschlel A P. Victoria R L, Saito S M T and Matsui E. 1982. 15N as a tool in biological nitrogen fixation in soil. In: P H Graham and S C Harris, eds. Biological nitrogen fixation technology for tropical agriculture. CIAT, Cali, Colombia. pp. 575-592.
Viriginia R A. 1980. Natural abundance of nitrogen-15 in selected ecosystems. Ph.D. Thesis, University of California, Davis. Diss. Abstr. Intern. 41.
Wagner G H and Zapata F. 1972. Field evaluation of reference crops in the study of nitrogen fixation by legumes using isotope techinques. Agron. J. 74: 607-612.
Walker T W. Adams A F R and Orchiston H D. 1956. Fate of labelled nitrate and ammonium nitrate when applied to grass and clover grown separately and together. Soil. Sci. 81: 339-351.
Wellman R P. Cook E D and Krouse H R. 1968. Nitrogen-15: Microbial alteration of abundance. Science 161: 269270.
West C P and Wedin W F. 1985. Dinitrogen fixation in alfalfa orchardgrass pastures. Agron. J. 77: 89-94.
Williams W A, Jones M B and Delwiche C C. 1977. Clover N-fixation measurement by total-N difference and 15N A-values in lysimeters. Agron. J. 69: 1023-1024.
Witty J F. 1983. Estimating N2-fixation in the field using 15N-labelled fertilizer: some problems and solutions. Soil Biol. Biochem. 15: 631-639.
Witty J F and Day J M. 1978. Use of 15N2 in evaluating asymbiotic N2 fixation. In: Isotopes in biological dinitrogen fixation. IAEA, Vienna, Austria. ST1/PUB/478. pp. 135-150.
Witty J F and Ritz K. 1984. Slow-release 15N fertilizer formulations to measure N2-fixation by isotope dilution. Soil Biol. Biochem. 16: 657-661.
Yamanaka K and Holl F B. 1984. Effects of N and seeding rate on grass-legume mixtures on coal mine spoils. Biomass production, soil factors, and N2 fixation. Agron. J. 76: 895-900.
![]()
![]()
![]()
{kind=link}