![]()
![]()
![]()
Abstract
Introduction
Environment and methods
Results and discussion
Conclusion
Acknowledgements
References
R.L. McCown1, W.H. Winter2, M.H. Andrew2, R.K. Jones3 and The Late D.C.I. Peake2
CSIRO Australia, Division of Tropical Crops and Pastures
1Davies Laboratory, Townsville, Queensland 4814.
2Darwin Laboratories, Darwin, Northern Territory 5789.
3ACIAR/CSIRO Project, c/o Australian High Commission, Nairobi, Kenya.
The notion of using sown pasture leys in rotation with annual crops to maintain soil fertility, reduce erosion and improve animal production has a long history in agricultural research in the savanna zone of Africa. However, research has shown little advantage of sown grass leys over vegetated fallow. The question of whether a legume ley would better provide the desired benefits has remained largely unanswered because, until recently, no tropical legumes that were sufficiently productive in the semi-arid tropics were available. This paper reports the findings from 4 years of a pilot study at Katherine, N.T., of a system in which cattle graze natural pastures in the wet season and croplands in the dry season. Croplands consist of one-third maize or sorghum and two-thirds pasture legume (2:1 rotation). The main objectives were to quantify (1) the contribution of grazed legume leys to the soil N supply of a succeeding cereal crop, (2) the benefits of legume leys to cattle growth and (3) the ability of legume pastures to withstand invasion of nitrophilous weeds. Three legumes, Stylosanthes hamata cv. Verano, Alysicarpus vaginalis, and Centrosema pascuorum, were compared.
In the one year that facilitated a field bioassay of N contribution, it is estimated the legume ley provided the equivalent of about 40 kg of fertilizer N/ha.
Steers on croplands in the dry season gained an average of 0.63 kg/day. Annual gains over 3 years averaged 123 kg, 30 kg more than cattle on natural pastures continuously.
Even within 2 years, it is clear that invasion of annual grass can be a problem, particularly in leys of A. vaginalis. In the longer term, the general encroachment of an aggressive perennial grass is judged to be an even greater threat.
Priority areas for further research in evaluating the potential of this farming strategy are identified.
The early impetus for research on ley farming in the savanna zone of Africa came with appreciation of the especially rapid decline of soil productivity when crop production was intensified. Colonial British agriculturalists, in particular, saw 'alternate husbandry', which had served agriculture in Britain so well, as an obvious solution where grazing animals were already important and the economic prospects for chemical fertilizers very limited. Their research aim was to provide 'the basis for a new and constructive system of pastoral agriculture' (Ross and Bumpus, 1544, quoted by Whyte, 1944, p. 34).
In the absence of a well adapted legume species, their attention focused on the benefits of sown grasses, but after five decades of research in East, West and southern Africa it can be safely concluded that grass leys have shown no advantage to crop yields over natural vegetated fallows (Webster and Wilson, 1980, p. 373). Contributions to nutrient supply to the crop phase are, at best, modest and short-lived (Barnes 1981: Stobbs 1969), and soil structure is improved for only one season on light textured soils (Wilkinson, 1975) and for little more than 2 years on heavy soils (Stephens, 1967). The persistent pursuit of a grass ley system for the tropics is understandable since it was not until the 1960s, 30 years after the start of the grass ley work in Africa, that British workers concluded that the main benefit of a temperate pasture ley to the succeeding crop is the nitrogen supplied (Cook 1967, p. 413), and that the legume component of the ley is crucial.
During the 1970s, well adapted pasture legumes for the savanna zone became available, and interest in tropical legume leys was renewed (Jones and Wild, 1975, p. 217; McCown et al. 1979; Webster and Wilson, 1980, p. 374). In 1978, research commenced at Katherine, Australia, to test the feasibility of a no-till, legume-ley system for the semi-arid tropics. The rationale, research strategy and preliminary results of the study of a number of components are reported by McCown et al (1985). This paper reports findings from the only experiment that was of sufficiently large scale to allow realistic grazing management. Thus it provides the only animal production information available from our work to date, as well as the only opportunity to observe the system working as a whole.
The objectives of the experiment were to measure (1) the contribution of grazed legume leys to the N supply for a cereal crop in rotation; (2) the benefits of legume leys to cattle growth, and (3) the ability of legume pastures to withstand invasion of nitrophilous weeds, particulary grasses, and to regenerate.
The physical environment for agriculture
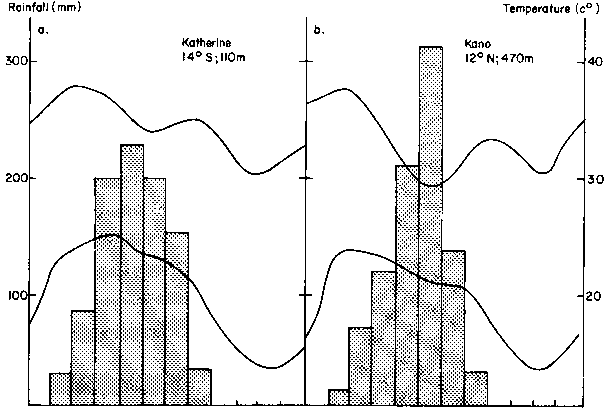
The climate and soils of the region around Katherine, N.T., Australia, have been described by Williams et al (1985). Katherine lies at 14° S at an elevation of 108 m. It has a mean annual rainfall of 930 mm, almost all of which falls during a hot summer (Figure 1a). Comparison with Figure 1b shows the close similarity of rainfall and temperature regimes to those of Kano, Nigeria. Experiments have shown that the climate is suitable for growing crops, including grain sorghum, maize, bulrush millet, cowpeas, soyabean, mung beans and peanuts, the main constraint being a risk of water deficits either early or late in the season. Rainfall erosivity is high and soil erosion is a major threat to agricultural productivity.
Beef cattle production on natural pastures is seriously constrained by the long dry season and low soil fertility. Winter rainfall events sufficient to benefit cattle nutrition by new grass production occur in many fewer years than in other areas of the Australian semi-arid tropics (McCown, 1982). However, this reliable dryness of the dry season is an important asset with sown legume pastures. In only one year in nine is there sufficient rain to cause spoilage of legume 'standing hay' much before opening rains of the new growing season (McCown, unpublished).
The soils of agricultural importance are Oxic Paleustalfs (Rhodic luvisols) ranging from clay loams to sands. The available water range is low (7-10%) even when clay content is high (Williams et al, 1985). Invariably the soils are deficient in N and P and occasionally in S or Zn. They are apedal and prone to slake when wet and to crust when dry.
Treatments
The experimental area consisted of 180 ha of natural grass/Eucalyptus open woodland and 3.6 ha of cropland. Within the cropland, one year of cereal crop was rotated with 2 years of legume pasture. The crop sequences for a 4-year period are shown in Table 1. Each sequence was followed in three blocks, each of which was sown to one of three legume species; Stylosanthes hamata cv. Verano, Alysicarpus vaginalis (Martin and Torssell 1974), and Centrosema pascuorum CPI 55697 (Clements et al, 1983). Neither the legume treatments nor the natural pastures were replicated. In the dry season, 12 steers were transferred from the natural pasture block to croplands where they had access to two-thirds of the area as dry legume and one-third as cereal residue. Six steers remained on natural pasture. Cattle in the croplands rejoined those on natural pastures following rainfall sufficient to initiate new growth on the natural pastures.
Table 1. Identification of crop sequences and the schedule of component crops and leys.
|
Crop sequence code |
1981/82 |
1982/83 |
1983/84 |
1984/85 |
|
A |
Maize |
Ley 1 |
Ley 2 |
Maize |
|
B |
Ley 1 |
Sorghum |
Ley 1 |
Maize |
|
C |
Ley 1 |
Ley 2 |
Maize |
Maize |
A further treatment split was that of the grain crop areas into two: in one half all weeds (including legumes) were controlled (IC0) and in the other grass weeds were controlled but pasture legumes allowed to form an understory or intercrop (IC+).
In summary, the croplands area was divided as follows:
|
Three legume species (grazed separately) |
3.6/3 = 1.2 ha |
|
Three rotation phases (toy 1, Ley 2, Crop) |
1.2/3 = 0.4 ha |
|
Two crop mixtures (+ Intercrop, No Intercrop) |
0.4/2 = 0.2 ha |
Pasture management and measurement
Legume pastures in each crop sequence (Table 1) were established initially with full cultivation, and thereafter were expected to regenerate naturally. Seeding rates were 30, 20 and 50 kg/ha of Verano stylo, Alysicarpus and Centrosema, respectively. Following the crop phase in sequence B (Table 1), density of Centrosema seedlings of the new pasture phase was inadequate: areas were scarificed lightly and seed broadcast at 26 kg/ha.
Grass was controlled in initial establishment with Treflan at 2.8 kg a.i./ha, and two weed species very much taller than the pasture legumes were controlled subsequently with glyphosate applied with a rope-wick boom.
The strategy for fertilization of the croplands was to keep all nutrients non-limiting. The only elements ever shown to be deficient in the region are N. P. S and Zn, with deficiencies of the last two occurring only occasionally. Prior to the experiment the area had received a large quantity of single superphosphate (Table 2). It was judged that the residual effects of this plus a further 200 kg of single superphosphate/ha in 1981/82 obviated the need for further applications on the pasture in 1982/83 and 1983/84. Similarly, residual effects of K and micronutrients from before the experiment and from crop phases in the experiment were judged sufficient for the pasture phases (Table 2).
Table 2. Fertilizer applied (kg/ha) to the croplands area.
|
Period |
Single superphosphate |
K |
Zn, Cu, Mo |
N |
|||
|
1978-81 |
1010 |
90 |
+ |
||||
|
|
Ley |
Crop |
Ley |
Crop |
Ley |
Crop |
Crop |
|
1981/82 |
200 |
200 |
0 |
50 |
+ |
+ |
35 |
|
1982/83 |
0 |
300 |
0 |
50 |
0 |
+ |
V++ |
|
1983/84 |
0 |
400 |
0 |
50 |
0 |
+ |
V |
|
1984/85 |
0 |
300 |
0 |
50 |
0 |
+ |
V |
+ Indicates 10 kg/ha each of copper and zinc sulphates and 0.2 kg/ha sodium molybdate; all fertilizer was broadcast except for superphosphate on crops in years 1981/82, 1982/83, and 1984/85.
++ Variable N rates on subplots; no N on main area.
Pasture vegetation in the croplands was sampled at three times. In late April, near the end of the main green season (as identified in Figure 3, below), the yield and composition of the pasture vegetation (including crop, weed and intercrop) were measured using the BOTANAL technique (Hargreaves and Kerr, 1978; Tothill et al, 1978). Pastures were sampled a second time at the start of the main dry season (as identified in Figure 3, below), when cattle were admitted, and a third time at the end of the main dry season, when cattle were removed. On these occasions, ten 0.5 m quadrats were harvested in each ley area of the three legume treatments.
The natural pasture area was typical of vegetation of the region. Each half of the area was burned in alternate years during the main dry season.
Crop management and measurement
Although in some years there is substantial rainfall in November and early December, its reliability is too low for planting before mid-December. Early rains result in regeneration of pasture species which yield 1 to 3 t DM/ha by crop planting time in most years. Crops were planted without prior tillage into pasture killed with glyphosate sprayed 1-2 weeks earlier at 2 1/ha. Maize was sown in 1981/82 and 1983/84; an early-maturing grain sorghum had to be substituted in 1982/83 due to the very late arrival of planting rains. Rows were 75 cm apart. Maize populations were 50-60,000/ha and sorghum 100,000/ha.
Fertilizer applications are shown in Table 2. Nitrogen (ammonium nitrate) was banded on the surface. In 1983/84 and 1984/85, plots within the main crop received a range of N rates. In each legume species treatment, rates of 0, 25, 50, 75, and 125 kg N/ha were replicated twice on plots 15 m x 4 rows.
At planting, the 'no intercrop, treatment was sprayed with 1.8 1 Dual-atrazine/ha to control both grasses and legumes. The 'intercrop' treatment received only Dual at 2.0 1/ha.
Grain yields reported (1983/84, 1984/85) were measured on 13 m lengths of the centre two rows of the N-treatment plots. Grain was oven dried and yields expressed on a 14% moisture basis. Stover dry weight was measured at the time of the second pasture sampling. Six 2.25 m2 quadrats were harvested in each of the six 0.2 ha crop areas.
Cattle management
Block licks containing NaCl, P. S and urea were available to cattle on natural pasture; the same mix minus urea was available to animals grazing croplands. Cattle were sprayed for tick control six times per annum. Cattle were returned from croplands to natural pasture areas when sufficient rain had fallen to produce substantial new growth on the burned area, i.e. 30-40 mm of rainfall.
At the start of each ley grazing period there were four animals in each ley pasture cell (two 1.5 years old and two 2.5 years old) and four animals grazing natural pasture (same age mixture). At the end of this period the older animals were replaced by yearlings. The reported growth of the ley/natural pasture group during the main green season is for the animals remaining in the trial only.
Cattle were weighed without fasting on the occasions indicated in Figure 3. Observations were made on grazing behaviour with regard to pasture and stover areas for one day every 2 weeks. Note was taken of the location of each steer and what it appeared to be eating at 15 minute intervals between 0730 and 1730.
Crop production
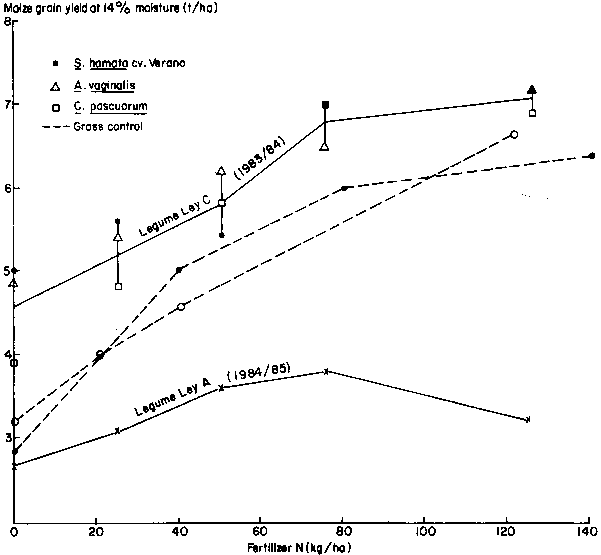
In only 2 years of the four could the effect of the 2-year ley on a succeeding crop be assessed (Table 1, sequences A and C). The 1984/85 season was unusually dry and yields were too low for the test crop bioassay for available soil N to be very informative (Figure 2). In 1983/84, water supply was ample, maximum yields were high and yields tended to increase linearly with fertilizer applications up to about 100 kg N/ha (Figure 2). To aid estimation of the N contribution of the legumes to the crop, data of two control grass treatments from other experiments have been included in Figure 2. Soil type and maize variety were the same as in the main experiment, and although grown in different years, maximum maize yields at the highest N rates were sufficiently similar (6.5-7 t/ha) to provide a helpful comparison with the effects of the legume leys.
An approximation of the fertilizer value of the legume ley can be obtained from Figure 2 as the amount of N that was needed on maize following grass to yield the same as maize following legume ley but without N fertilizer. This was about 40 kg N/ha for the 1983/84 crop. The pasture that provided this effect had a dry-matter yield of 6.6 t/ha in the first year, 90% of which was legume (toy 1, sequence C); Ley 2 (second year) yielded 5.8 t/ha, 57% of which was legume (Table 3).
Differences in maize yield after Verano stylo and Alysicarpus leys were small (Figure 2). However, after Centrosema, maize yield without N fertilizer was about 20% less than after the other two legumes (Figure 2), even though yields of legume in the preceding Centrosema ley were as high or higher than in the other two leys (Table 3). If this difference is real, this is particularly puzzling since the N concentration in C. pascurorum is higher than in the other two legumes (data not shown).
Table 3. Yield and composition of leys sown to Stylosanthes hamata cv. Verano, Alysicarpus vaginalis, and Centrosema pascuorum in both first and second years in two crop-fey sequences (Table 2). (Fortes other than the three legumes are not reported and account for the deviation of totals from 100%.
|
Sown legume species |
|
Sequence C |
Sequence A |
||
|
Ley 1 |
Ley 2 |
Ley 1 |
Ley |
||
|
Stylosanthes |
Dry weight (t/ha) |
6.6 |
5.8 |
4.7 |
7.4 |
|
S. hamata (%) |
85 |
44 |
62 |
36 |
|
|
A. vaginalis(%) |
10 |
13 |
25 |
25 |
|
|
C. pascuorum (%) |
0 |
0 |
0 |
0 |
|
|
Grass (%) |
2 |
39 |
6 |
38 |
|
|
Alysicarpus |
Dry weight (t/ha) |
4.8 |
6.2 |
5.5 |
7.1 |
|
S. hamata (%) |
10 |
39 |
0 |
2 |
|
|
A. vaginalis (%) |
85 |
32 |
77 |
12 |
|
|
C. pascuorum (%) |
1 |
0 |
0 |
0 |
|
|
Grass (%) |
0 |
29 |
22 |
85 |
|
|
Centrosema |
Dry weight (t/ha) |
7.1 |
6.5 |
5.1 |
9.3 |
|
S. hamata (%) |
3 |
24 |
1 |
6 |
|
|
A. vaginalis (%) |
3 |
23 |
15 |
15 |
|
|
C. pascuorum (%) |
72 |
22 |
77 |
70 |
|
|
Grass (%) |
0 |
29 |
1 |
8 |
|
The results of these 2 years raise more questions than they answer. Clearly large differences in benefits can occur, but what is the frequency distribution of benefits among years? Benefits to unfertilized maize yields in the 'good' season of 1983/84 were about two-thirds those of a one-year, ungrazed, pure legume sward reported by McCown et al, (1985) and about half of the highest we have measured following pure ungrazed swards. To what extent are differences due to effects of grazing or due to effects of grass being present? In the next phase of this study, a much larger sample of years will be obtained. At the same time, quantification of rates of mineralisation, degree of immobilisation due to the presence of grass and loss rates as affected by soil type and water climate will be monitored in closely related studies.
Animal production
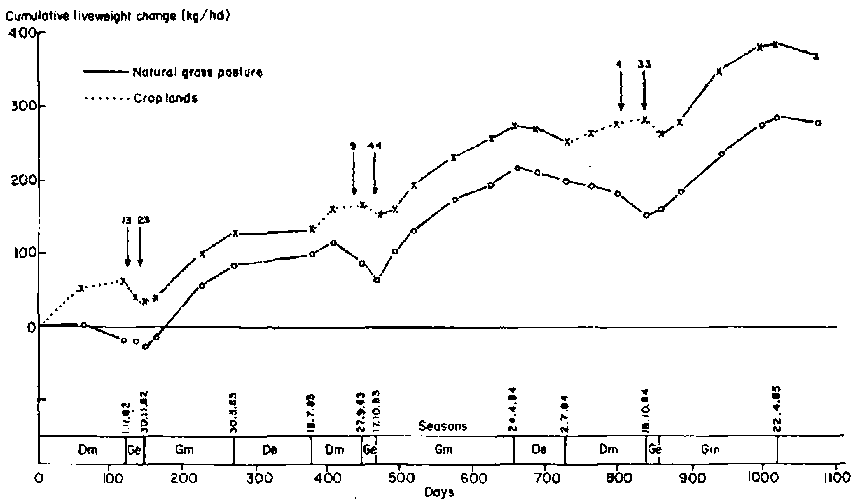
Trends in cummulative liveweight change of cattle for a 3-year period averaged over the three legume species are shown in Figure 3. Animals grazing continuously on natural pasture gained an average of 93 kg per annum, whereas those on croplands during the dry season averaged 123 kg. To assist analysis of these differences, years in Figure 3 have been divided into seasons: main dry (Dm), when animals were grazing croplands; early green (Ge), a period of erratic rainstorms when cattle on croplands return to natural pastures; main green (Gm), the period of weight gain on natural pastures; and early dry (De), a non-gain period on natural pasture prior to availability of croplands.
Cattle gained an average of 456 g/head per day when grazing croplands in the main dry season, 705 g more than those that remained on grass pasture (Table 4). Part of the advantage gained by cattle in the croplands during the main dry season was lost in other seasons. In 2 of the 3 years in which cattle remained on the legume leys following more than a few millimeteres of rain in the early green season, weight loss was rapid (Figure 3). This was presumably due to intake suppression as a result of moulding (McCown et al, 1981). Losses that occurred in animals on the natural pastures were probably due to reduction in gut fill with the appearance of first green grass regrowth (McLean et al, 1983). Further weight advantages gained by cattle in the croplands were lost on natural pastures during the main green and early dry seasons (Table 4, Figure 3).
Table 4. Rates of liveweight change of cattle grazing (a) natural pastures the entire year and (b) native pastures in the green season and legume leys in the dry season (averages of three years).
|
|
Duration (days) |
Continuous natural pasture |
Natural pastures+ leys |
|
Liveweight change (g/hd/day) |
|||
| Dry season | |||
|
Early |
90 |
-67 |
-171 |
|
Main |
99 |
-249 |
+456 |
|
Green season |
|
|
|
|
Early |
22 |
-393 |
-964 |
|
Main |
159 |
+827 |
+733 |
Liveweight gains of animals grazing the three legume species treatments did not differ greatly except in the main dry season in 1984, when rate of gain of animals grazing Alysicarpus was much less than that of animals grazing the other two legumes (Table 5). This low rate of gain was associated with the lowest total legume (48%)/highest grass (52%) pasture composition in the experiment (Table 6). In the one season (1982) when the purity of the sown species was high in all paddocks (Table 6), rates of gain differed by less than 3% (Table 5). In subsequent years, interpretation in terms of differences in animal production between legume species is hampered by the mixing of legumes that had occurred (Table 3).
Table 5. Liveweight changes during the main dry season on four pastures.
|
|
Year |
||
|
1982 |
1983 |
1984 |
|
|
|
Liveweight change (g/hd/day) |
||
|
Verano ley |
+448 |
+559 |
+443 |
|
Alysicarpus ley |
+449 |
+451 |
+235 |
|
Centro ley |
+459 |
+541 |
+467 |
|
Natural grass |
-168 |
-143 |
-238 |
The amounts of pasture dry matter available at the beginning of the main dry season ranged from 3.8 to 5.6 t/ha, comprised in most cases of more than 70% legume (Table 6). About 2 t/ha generally remained at the end of the period. Rate of disappearance per day was 7.3, 9.2, and 8.6 kg for the 3 years.
The nutritional contribution of stover would have been modest. Crop on one-third of the grazed area provided 0.30.5 t of stover per animal, much of which, although unmeasured, was not eaten. Quality of maize stover is normally low and the normally more valuable sorghum did not ratoon significantly in 1983. Time spent grazing maize stover declined from 13% at the beginning to 2% by the end of the main dry season. In 1983, time spent on sorghum stover declined with time from 15 to 9%.
Table 6. Yields and composition of croplands forage when cattle entered and when they left in three main dry seasons (Figure 3).
|
|
Stover DM (t/ha) |
Initial pasture DM (t/ha) |
Proportion of legume (%) |
Residual pasture DM (t/ha) |
|
1982 |
|
|
|
|
|
S. hamata |
|
5.0 |
93 |
1.7 |
|
A. vaginalis |
|
3.8 |
90 |
1.6 |
|
C. pascuorum |
|
5.2 |
81 |
2.1 |
|
Mean |
1.5 |
4.6 |
88 |
1.8 |
|
1983 |
|
|
|
|
|
S. hamata |
3.8 |
74 |
2.1 |
|
|
A. vaginalis |
4.3 |
77 |
1.8 |
|
|
C. pascuorum |
4.3 |
78 |
2.0 |
|
|
Mean |
1.0 |
4.1 |
76 |
1.9 |
|
1984 |
|
|
|
|
|
S. hamata |
5.6 |
67 |
2.0 |
|
|
A. vaginalis |
5.2 |
48 |
2.0 |
|
|
C. pascuorum |
1.0 |
5.6 |
79 |
2.9 |
|
Mean |
|
5.4 |
65 |
2.3 |
Botanical stability
Annual grass invasion: The third major objective of this study relates to the threat of invasion of legume leys by grass. Under conditions of low soil P and with grazing during the wet season (when grass is grazed preferentially), legume pastures in this environment have retained a high degree of purity for many years (Winter et al, 1985). However, under conditions of high P input and grazing deferred until the dry season, annual grass invaded and displaced Townsville stylo after a few years (Gillard and Fisher, 1978). Our hypothesis in the present experiment was that by using legume species that are more competitive than Townsville stylo, and with ley durations of only 2 years before cropping with nitrophilous crops which deplete available soil nitrogen levels (and when grass is 'eradicated' with herbicides), grass content of leys could be kept to a tolerable level.
Table 3 shows the changes in botanical composition from the first to the second year of ley in two crop sequences (Table 1). Ley 1, sequence C was the original pasture in which every effort was made to ensure high purity of the sown legume species. Legume contamination was only 10% and grass no more than 2%. By the second year, Verano stylo in the stylosanthes treatment had dropped from 85% to 44% and grass had increased from 2 to 39%. The proportion of alysicarpus in the alysicarpus treatment dropped from 85% to 32% due mainly to an increase in grass from nil to 28% and, of less consequence, an increase in Verano stylo from 10 to 39%. The proportion of Centrosema dropped from 72 to 22% due to an increase in grass from nil to 29% and invasion of both other legumes. Ley 2 in sequence C was succeeded by the maize crop in 1984 whose N response is shown in Figure 2.
Ley 1 of sequence A (Table 1) was sown without herbicides, and purity was lower than in sequence C. Initial foreign legume content, although quite high in stylosanthes and centrosema treatments, did not increase in Ley 2. Grass invasion varied dramatically among legume treatments. There was remarkably little invasion into centrosema. The pattern in stylosanthes was similar to that in sequence C. However in alysicarpus, grass content in Ley 1 was 22% which increased to 85% in Ley 2.
Legume regeneration: An opportunity to compare the abilities of the three legumes to re-establish pastures in the year following the crop occurred in sequence B (Table 2). In 1982 leys of the three species, over 80% of the leys consisted of the species sown (Table 7). Total dry-matter yields exceeded 5.5 t (Table 6). In the crop in 1983, Alysicarpus comprised a high proportion of both weed and intercrop vegetation (Table 7), even in stylosanthes and centrosema treatments. In 1984, ley pastures of alysicarpus and Verano established swards with acceptable amounts of the nominal legume without assistance. Centrosema failed to reestablish and the block was scarified lightly and re-sown; despite this, centrosema comprised only 12% of the yield at the end of the growing season (Table 7).
C. pascuorum is not easy to establish on bare soil. In contrast to the other two species, which have small seeds, the large radicle of this large-seeded species has difficulty penetrating even wet soil without counter-resistance on the seed by soil or mulch cover.
Perennial grass invasion: The perennial grass Urochloa mozambicensis cv. Nixon is very well adapted to this climate and, where it has been introduced, rapidly becomes a dominant component of permanent pastures where P fertilizer inputs are adequate. In this experiment, vegetation of border areas and fence lines was dominated by this species and received no special control measures. Although U. mozambicensis had been gradually increasing overall, the grass dominance, and particularly that of U. mozambicensis, increased dramatically between the 1984 and 1985 seasons (Table 8). Even allowing for the fact that the balance between grass and legume normally shifts toward legume as a season progresses, both the 93% total grass content of the new Ley 2 and the 56% U. mozambicensis content of half of Ley 1 are alarming. The reason for the very much higher U. mozambicensis content in the 'no intercrop' treatment is not clear, but it is not accounted for by differences in U. mozambicensis content in the previous ley phase (data not shown).
Table 7. Botanical changes in the pasture component of crop-fey sequence B. (Fortes other than the legumes indicated are not reported and account for the deviation of totals from 100%).
|
Sown legume species
|
Actual components
|
Percent by weight |
||
|
Ley |
Crop |
Ley |
||
|
1982 |
1983 |
1984 |
||
|
Stylosanthes |
S. hamata |
84 |
11 |
36 |
|
A. vaginalis |
10 |
78 |
34 |
|
|
C. pascuorum |
0 |
0 |
0 |
|
|
Grass |
1 |
7 |
27 |
|
|
Alysicarpus |
S. hamata |
16 |
0 |
12 |
|
A. vaginalis |
80 |
91 |
64 |
|
|
C. pascuorum |
0 |
0 |
0 |
|
|
Centrosema
|
S. hamata |
2 |
1 |
22 |
|
A. vaginalis |
1 |
32 |
30 |
|
|
C. pascuorum |
83 |
10 |
12 |
|
|
Grass |
0 |
10 |
36 |
|
* Oversown in response to failure to re-establish naturally.
Table 8. Total dry matter and botanical composition of seedling vegetation early in the 1984/85 season (12 Dec 1984).
|
Treatment in 1984: |
Crop/IC |
Crop/No IC |
Ley 1 |
|
1985: |
Ley 1 |
Ley 1 |
Ley 2 |
|
Total DM (t/ha) |
1.4 |
1.5 |
2.2 |
|
Legume % |
45 |
34 |
2 |
|
Annual grass % |
42 |
8 |
93 |
|
U. mozambicencis % |
8 |
56 |
3 |
* Intercrop of pasture vegetation.
In the present study, crop height was dramatically depressed in patches that had been dominated by U. mozambicensis in the previous fey. This was presumably due to a much greater immobilization of soil N. Considering the apparent rapid increase in the proportion of U. mozambicensis, the implications or. system performance and the difficulty of selectively eliminating this species, it was decided to terminate the study. This was done in time to plant a uniform crop of maize in the 1985 season, accompanied by an intensive weed control program. It is planned to recommence the study when the U. mozambicensis problem is under control, and to manage the area in the future to minimise access of this species.
For a system in which a pasture or forage crop displaces a grain crop to be economically feasible in either Australia or Africa the synergistic benefits have to be substantial. A gross comparison of production of the ley system and that of continuous cropping and permanent grazing is made in Table 9. In spite of productivity benefits to the crop, total maize grain yields in the ley system are estimated to be about half that of a continuous cropping system. Cattle liveweight gains were 360 kg (22%) higher than on continuous natural pasture grazing.
At this stage, simplistic economic analysis of the differences is likely to be misleading. In the first place, there is evidence that in neither Africa or Australia would such ley pastures be used for fattening cattle. In northern Nigeria, Fulani pastoralists tend to use Verano stylo fodder banks in the dry season to ensure survival of vulnerable animals rather than for production (von Kaufmann, personal communication). In Australia preferred use might be for weaners - to ensure good growth rates during weaning and training in the confinement of small paddocks. In both cases the monetary benefits are substantial, but more difficult to quantify than liveweight of slaughter cattle.
Table 9. Comparison of the total annual production of three systems.
|
|
Maize grain |
Cattle |
||
|
Area |
Yield |
Area |
Wt gain |
|
|
(ha) |
(t) |
(ha) |
(kg) |
|
|
Continuous cropping |
3.6 |
10.8* |
- |
- |
|
Continuous natural pasture grazing (18 steers) |
- |
- |
180+? |
1670 |
|
2:1 fey-crop rotation |
1.2 |
5.5 |
180+3.6 |
2030 |
* Yield of maize following a crop and with no nitrogen fertilizer assumed to be 3 t/ha.
In the Australian context, where it can be presumed that it is unprofitable not to remedy non-nitrogen nutrient deficiencies in cropland, depreciation of the legume benefits by grass invasion of well fertilized leys may be the most serious threat to success of this hypothetical system. (In the next phase of this study much greater attention will be paid to the dynamics of weeds.)
In the African context, problems of restricted availability and high costs of supplying non-nitrogen fertilizer seem inescapable, and the implications of phosphorus deficiencies on both crop and animal enterprises of the legume ley system need to be quantified. With regard to crops, both legume N production and the response of the cereal crop to the increased N supply are sensitive to
P supply. Research on these interactions with P supply and the further interaction with legume species was recently started at Katherine, and collaborative research is planned in Kenya and northern Nigeria. With regard to animals, the degree to which P fed as mineral supplements can compensate for low fertilization of legume pastures is currently under study at Katherine and collaborative research is planned in northern Nigeria.
The technical assistance of N.P. Dalgleish, J.J. Hodgkinson, and J.P. Corfield is gratefully acknowledged. In the third year, this research benefited from financial assistance from the Australian Centre for International Agricultural Research.
Barnes D L. 1981. Residual effects of grass leys on the productivity of sandy granite-derived soils. 2. Grazed leys. Zimbabwe J. Agric. Res. 19: 69-82.
Clements R J. Williams R J. Grof B and Hacker J B. 1983. Centrosema. In: R L Burt, P P Rotar, J L Walker and M W Silvey, (eds). The role of centrosema, desmodium and stylosanthes in improving tropical pastures. Westview Press, Boulder, USA.
Cook G W. 1967. The control of soil fertility. Crosby Lockwood, London.
Gillard P and Fisher M J. 1978. The ecology of Townsville stylo-based pastures in northern Australia. In: J R Wilson, (ed). Plant relations in pastures. CSIRO, Melbourne, Australia.
Hargreaves J N G and Kerr J D. 1978. A comprehensive sampling and computing procedure for estimating pasture yield and composition. II. Computational package. Tropical Agronomy Technical Memorandum No. 9. Div. Tropical Crops and Pastures, CSIRO, Australia.
Jones M J and Wild A. 1975. Soils of the West African savanna: the maintenance and improvement of their fertility. Technical Communication No. 55. Commonwealth Bureau of Soils, Harpenden, U.K.
McCown R L. 1982. The climatic potential for beef cattle production in tropical Australia. Part IV. Variation in seasonal and annual productivity. Agric. Systems. 8: 3-15.
McCown R L, Haaland C and de Haan C. 1979. The interaction between cultivation and livestock production in semiarid Africa. In: Hall A E, Cannell G H and Lawton H W. (eds). Ecological studies, Vol. 34. Agriculture in semiarid environments. Springer-Verlag, Berlin.
McCown R L, Wall B H and Harrison P G. 1981. The influence of weather on the quality of tropical legume pasture during the dry season in northern Australia. I. Trends in sward structure and moulding of standing hay at three locations. Aust. J. Agric. Res. 32: 575-87.
McCown R L, Jones R K and Peake D C I. 1985. Evaluation of a no-till, tropical legume ley-farming strategy. In: Muchow R.C, (ed). Agro-research for the semi-arid tropics: North-west Australia. University of Queensland Press, St Lucia, Australia.
McLean R W. McCown R L, Little D A, Winter W H and Dance R A. 1983. An analysis of cattle live-weight changes on tropical grass pasture during the dry and early wet seasons in northern Australia. 1. The nature of weight changes. J. Agric. Sci. (Camb). 101: 17-24.
Martin M H and Torssell B W R. 1974. Buffalo clover (Alysicarpus vaginalis (L) (DC.)): A pasture legume in Northern Australia. J. Aust. Inst. Agric. Sco. 40:232-4.
Stephans D. 1967. Effects of grass fallow treatments in restoring fertility of Buganda clay loam in south Uganda. J. Agric. Sci. (Camb.) 68: 391-403.
Stobbs T H. 1969. The effect of grazing resting land upon subsequent arable crop yields. E. Afr. Agric. For. J. 35:28-32.
Tothill J C, Hargreaves J N G and Jones R M. 1978. BOTANAL: A comprehensive sampling technique for estimating pasture yield and composition. 1. Field sampling. Tropical Agronomy Technical Memorandum No. 8. Div. Tropical Crops and Pastures, CSIRO, Australia.
Webster C C and Wilson P N. 1980. Agriculture in the tropics. Longmans, London.
Whyte R O. 1944. Trends in different countries and regions. In: Alternate Husbandry. Imperial Agricultural Bureaux Joint Publ. No. 6, Gt. Britain.
Wilkinson G E. 1975. Effect of grass fallow rotations on the infiltration of water into a savanna zone of Northern Nigeria. Trop. Agric. (Trinidad) 52: 99-103.
Williams J. Day K J. Isbell R F and Reddy S J. 1985. Soils and climate. In: Muchow R C, (ed). Agro-research for the semi-arid tropics: North-west Australia. University of Queensland Press, St Lucia, Australia.
Winter W H. McCosker T H. Pratchett D and Austin J D A. 1985. Intensification of beef production. In: Muchow R C (ed.) Agro-research for the semi-arid tropics: Northwest Australia. University of Queensland Press, St Lucia, Australia.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}