![]()
![]()
![]()
In order to understand livestock water needs and how water dependence varies in different circumstances, it is necessary to look at body water functions in livestock, starting with the way water is stored and distributed in the animal.
Under semi-arid conditions and a daily watering regime, indigenous livestock in Africa use, or turn over, between 5 and 30% of their body water pool (defined in section 2.2) daily, depending on animal species and season (King, 1979). This water is used for two main functions; intermediary metabolism and cooling.
Intermediary metabolism includes all the biochemical reactions and interconversions that take place in the animal. Water acts as a solvent of absorbed particles, including gases, conveying them to and from their sites of metabolism. The resultant solutions osmotically control the distribution of fluids within the body compartments, with the aid of various feedback regulating mechanisms. This aspect of the subject is beyond the scope of this report, but for those who wish to pursue it further the main components of the intermediary metabolic system are described by Siebert (1973).
In most herbivores in sub-Saharan Africa, the major function of water is to dissipate internal or absorbed heat by evaporative cooling through sweating or panting.
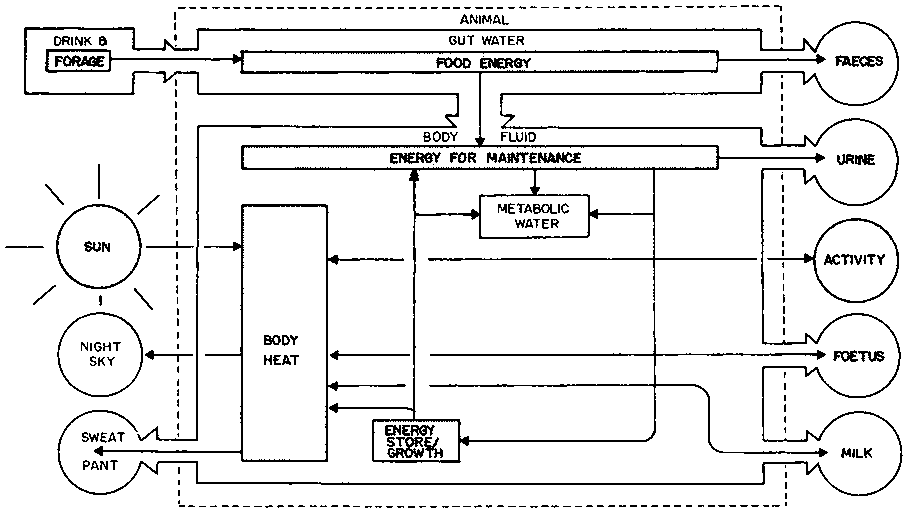
The flows of water and energy through the animal body have been simplified and combined in Figure 2. Most of these flows can be quantified, and an energy and water budget measured. For example, in zebu cattle in the Kenya highlands during the day, the main heat exchanges have been shown to be absorption of shortwave solar radiation and emission of longwave radiation by the coat (Table 5). About 1% of the energy ex changed is heat which is absorbed and stored during the day and dissipated at night. In the tropical lowlands, the environmental heat load is higher and evaporative heat loss more important.
About 99% of all molecules in the body are water, which forms about 70% of the bodyweight of a tropical ruminant (Macfarlane and Howard, 1972). Its distribution through the body water compartments is fairly constant if the alimentary tract and fat content of the animal are excluded: about 45% of bodyweight is intracellular water and 25% is extracellular, divided between plasma (5%) and interstitial fluids (20%).
Table 5. Energy budget of a zebu steer from 09.00 to 15.00h in a near-natural environment at 1430 m altitude in Kenya.
|
Heat exchange |
% of net gain |
% of net loss |
|
Absorption of shortwave solar radiation |
71 |
|
|
Metabolic heat production |
29 |
|
|
Longwave re-radiation from coat |
|
57 |
|
Sweating |
|
21 |
|
Conduction and convection from skin |
|
16 |
|
Respiratory evaporation |
|
5 |
|
Total |
100 |
99 |
Source: Finch (1976).
Figure 2. Water and energy flow through an ungulate in Africa.
The total body water pool is all the water in the animal including the alimentary tract, which has a large volume, particularly in ruminants. The moisture content of the reticulo-rumen seldom falls below 85%, even on a dry diet (Macfarlane, 1971). Based on this moisture content, the average contribution of reticulo-rumen water to total body water has been calculated for a variety of African ruminants (Table 6), classified according to feeding habits (Hofmann, 1973). The volume of rumen contents may vary by nearly 100% between the wet and dry season or even on alternate days in livestock on a 2-day watering regime, when sheep drink 18% of their total body water pool, goats 20%, zebu cattle 14% and camels 10% (King, 1979). Larger fluctuations are associated with enforced dehydration.
The proportion of body water in the other carcass components of a ruminant are given in Table 7, using as an example a 2-year-old Boran steer fed a dry maintenance ration. It can be seen that the components with the lowest water content are bone and fat, although the values are higher than the 20% and 6-20% respectively reported by Brody (1945), presumably because of a higher proportion of connective tissue. Bone is relatively inert, but fat represents the main energy pool of the animal, with a value of 39.3 kJ.g-1 compared with protein at 18.0 kJ.g-1 and carbohydrate at 17.6 kJ.g-1 (Lusk, 1931). Thus the energy value of the carcass of most domestic animals has been derived from analysis based primarily on the fat content (Callow, 1947). Because of its low water content an increase in the proportion of body fat is often associated with a decrease in percentage body water pool. At birth, the fat content of the body is negligible and the body water pool is about 77% of bodyweight. As the animal matures, fat is deposited until it accounts for up to 35% of bodyweight in ruminants in temperate regions, while total body water can drop to 50%. It is difficult to measure the fat content of a live animal directly, but it has often been estimated indirectly from the tritium-predicted body water pool (e.g. Shumway et al, 1956; Searle, 1970). When this technique was applied to the zebu in Africa (King and Finch, 1982), the following regression was obtained:
y = 60.46 - 0.801x (r2 = 0.798, P <0.001)
where y equals dissectable body fat, and x equals tritium-predicted body water pool, all expressed as a percentage of bodyweight of zebu steers with a dissectable body fat of <10%. Although the inverse relationship was expected, it was not easy to explain in lean, starving animals in which the proportions of other components with very different water contents, such as bone and hide, were also changing. The use of such prediction equations outside the conditions in which they were derived is therefore unwise. As Webster (1978) has remarked, 'The quite common predictive use of correlations between marker dilutions and the contents of compartments they do not mark, is a thoroughly bad habit'. One can reduce the error by using the approach of Little and MacLean (1981), who noted that in castle and sheep the sum of total body water plus total body fat minus the DM content of the alimentary tract was virtually 80% of the fasted bodyweight.
The long held belief that there are sacs in the forestomachs of the camel specifically for storing water has been refuted by Schmidt-Nielsen (1965). Nevertheless, after drinking water moves two or three times more slowly from the reticulo-rumen to the blood in camels than it does in cattle (Siebert and Macfarlane, 1971). The release of ingested water through the kidney does not start until after 1 day in camels, compared with 12 h in Merino sheep, and 4 h in Bos taurus cattle (Macfarlane and Howard, 1972). The main purpose of this retention is to prevent hypotonic solutions passing into the bloodstream until salts have been added. Nevertheless the result is some degree of water storage, particularly in the more arid-adapted animals.
Table 6. Proportion of total body water in the reticulo-rumen of some African animals classified according to feeding habits.
|
Grazers |
Mixed feeders |
Selective concentrate feeders | |||
|
Species |
% |
Species |
% |
Species |
% |
|
African buffalo |
20 |
Haired sheep |
24 |
Grey duiker |
12 |
|
Wildebeest |
18 |
Haired goat |
24 |
Suni |
10 |
|
Zebu |
17 |
Eland |
15 |
Steenbok |
9 |
|
Oryx |
15 |
Thomson's gazelle |
15 |
Bushbuck |
9 |
|
Hartebeest |
14 |
Camel |
13 |
Dik-dik |
9 |
Source: Schmidt-Nielsen (1965); Hofmann (1973); King et al (1975); Kay et al (1980).
Table 7. Water content of carcass components of a 2-year-old Boran steer.
|
Carcass component |
|
Water content |
|
|
Weight (kg) |
% |
l |
|
|
Slaughter loss |
7.0 |
80a |
5.6 |
|
Blood |
7.2 |
81 |
5.8 |
|
Hide |
18.0 |
67 |
12.0 |
|
Horns |
- |
- |
6.9 |
|
Head |
13.0 |
54 |
|
|
Feet |
6.0 |
49 |
2.9 |
|
Heart, diaphragm, respiratory, urinary and genital tracts |
5.6 |
72 |
3.8 |
|
Urine and bile |
0.1 |
90a |
0.1 |
|
Liver |
2.4 |
72 |
1.7 |
|
Contents of forestomachs |
18.8 |
86 |
16.1 |
|
Contents of small intestine |
4.3 |
90 |
3.9 |
|
Contents of large intestine |
4.9 |
82 |
4.1 |
|
Gut wall |
10.9 |
80 |
8.8 |
|
Dissectable fat: internal |
3.4 |
40 |
1.4 |
|
Dissectable fat: subcutaneous, intermuscular, hump |
11.2 |
47 |
5.3 |
|
Butcher's bone: legs |
14.6 |
20 |
2.9 |
|
Butcher's bone: loin |
2.8 |
42 |
1.2 |
|
Butcher's bone: neck and thorax |
7.7 |
34 |
2.6 |
|
Carcass lean |
74.3 |
75 |
56.0 |
|
Kidney |
0.3 |
75 |
0.3 |
|
Dissection loss |
3.6 |
80a |
2.9 |
|
Total or average |
216.1 |
67 |
144.3 |
a Estimate
Source: J.M. King and V.A. Finch (unpublished).
Water also appears to be stored in the extracellular and intracellular fluid spaces in some ungulates under certain environmental conditions, judging by their tolerance of 15 - 20% dehydration without depression of appetite and lactation. For example, Shkolnik et al (1972) reported expanded plasma volumes in Bedouin goats associated with high tritiated water (TOH) spaces, although their figures of 85% TOH spaces may need downward correction. It has also been shown in cattle and buffaloes in the tropics that, as the water turnover rate increases in response to heat stress, the body water pool enlarges (Siebert and Macfarlane, 1969; Kamal and Sief, 1969).
These illustrations suggest that there is a need to re-examine the concept that a camel drinks 'for the past' and not 'for the future'. It certainly drinks to make up a deficit, but its intake may also include a provision for the future.
The oedematous condition is characterised by an excessive accumulation of fluid in tissue spaces. Fluid normally passes into these spaces from the arterial end of the capillaries, where the hydrostatic pressure of the blood is greater than its osmotic pressure. It will stay there if the position is not reversed at the venous end of the capillaries or if there is a blockage of lymphatic drainage (Schmidt-Nielsen, 1975).
A non-pathological oedematous condition is said to be achieved by camel traders, who allow a very thirsty camel to drink its fill after it has been given a large dose of salt. The resultant subcutaneous oedema can make a thin camel look temporarily well covered (Williamson arid Payne, 1978).
There are numerous cases of pathological oedema in African livestock, frequently associated with a reduced blood osmotic pressure due to hypoproteinaemia. For example, failure of amino-acid anabolism is associated with malnutrition and liver disease; continuous haemorrhage from the gastrointestinal tract, resulting in a loss of blood volume and erythrocytes as well as plasma protein, is caused by infestations of blood sucking nematodes, or by coccidia or salmonella; trypanosomes destroy erythrocytes. As a result, fluid drains to the dependent parts of the body, notably the submandibular space, brisket and abdomen. Oedema of the limbs, scrotum or vulva are additional features of trypanosomiasis in the horse (Blood and Henderson, 1960; BVA, 1976).
During starvation there comes a point when the energy reserves of the animal have become severely depleted and the rise in total body water changes from a physiological to a pathological condition, probably associated with a rise in extracellular fluid volume (Macfarlane et al, 1966c; Springell, 1968). The stage at which this occurs in the zebu must be shortly after the dissectable fat reserves have been exhausted, when its total body water pool is 70% of its liveweight (King and Finch, 1982). The clinical picture of oedema can be complicated by dehydration and, in acute cases, by peripheral circulatory failure and anaemic anoxia (Blood and Henderson, 1960).
Blockage of the lymphatic or venous drainage occurs following liver fibrosis and obstruction of the hepatic portal. For example, liver flukes cause ascites - an accumulation of fluid in the abdominal cavity. Cardiac insufficiency, due to endocarditis or pericarditis will also cause oedema. The resultant congestion of the lungs is a common terminal event (Blood and Henderson, 1960). In East Coast fever in cattle such congestion may also be due to a terminal anaphylactic reaction (A.D. Irvin, unpublished), which is another cause of oedema.
![]()
![]()
![]()
{kind=link}