![]()
![]()
![]()
2.1 HYDROGRAPHY
2.2 FAUNISTIC REGIMES
2.3 THE SHELF AND SLOPE, AREAL EXTENTIONS AND BOTTOM CONDITIONS
* This chapter is contributed by Dr. ph. H. Gade, Institute of Geophysics, University of Bergen.2.1.1 General physiographic conditions of the region
2.1.1.1 Topography
In the following discussion of the topography the survey area is divided in three coastal regions corresponding to the presentation of the hydrography of the waters. These regions are 1) Suriname, Guyana and eastern Trinidad-Tobago, 2) Caribbean waters from Trinidad-Tobago to Gulf of Venezuela and 3) Colombian waters from Guajira Peninsula to Gulf of Uraba.
Coastal waters off Suriname, Guyana and eastern Trinidad-Tobago
The general features of the topography of the region are evident from the charts showing the survey area. The continental shelf is here the widest of the discussed regions, ranging from 100 nm in the southernmost part (Maroni) to about 35 nm east of Trinidad. The shelf break lies at depths from 90 m to 100 m. Outside the shelf break the continental slope descends to depths of several thousand metres. The continental shelf has a fairly smooth topography with no recognised canyons or other submerged features. Just north of Tobago the shelf edge turns sharply westwards, with Tobago in its entirety on the continental shelf. Oceanographically important is the opening between Tobago and Trinidad, which is 19 nm wide and has an average depth of about 50 m. The sill depth is about 90 m.
Southeastern Caribbean waters from Tobago to Gulf of Venezuela
The width of the continental shelf increases gradually from 27 nm north of Trinidad to almost 60 nm north of the Arya Peninsula, as measured out to the 200 m depth line. With allowance for topographic features such as Blanquilla Island, Margarita Island, Araya Peninsula and Cariaco Trench, the shelf width remains fairly much the same as far west as Tortuga Island. From there on the shelf width decreases fairly sharply as the 200 m depth contour line and the coastline converge. Outside La Guaira the shelf width is down to about 5 nm decreasing further to the west to be virtually nonexistent shortly east of Golfo Triste. Here it widens out to about 25 nm from there on varying between 10 and 20 nm to Paraguana Peninsula. Off Paraguana Peninsula the continental shelf widens to include Aruba, leaving Gulf of Venezuela as a large shallow sea with depths of about 70 m in the open northerly parts. To the south the gulf bottom ascends gradually towards a break at about 30 m. North of Guajira Peninsula the shelf narrows again down to a width of about 10 nm.
Guajira Peninsula to Gulf of Uraba
In this part of the Colombian coast the continental shelf is fairly narrow. North of Guajira Peninsula the 200 m contour line is found about 10 nm off the coast. Westwards the shelf widens to a maximum of about 25 nm in the Riohacha area. Further west the shelf width diminishes again and practically vanishes off, but widens out to about 10 nm north of Barranquilla. Further south the shelf width varies from 10 to 30 nm, the latter also being the width outside Gulf of Uraba. The shelf break in the Colombian region is clearly more shallow than in the Venezuelan region, lying mostly in the 50-60 m depth range. The average shelf depth is correspondingly less.
2.1.1.2 Meteorology and climatology
Both from meteorological and oceanographic points of view the surveyed area maintains a rather stable climate on a short time scale basis, however, subjected to a fairly strong seasonal signal. This is due to the stability of the trade winds which blow steadily throughout 3/4 of the year. Only during the rainy season (August-November) does the weather exhibit a fair degree of variability, leading to heavy showers and thunderstorms, particularly in the easternmost part of the survey area, i.e. in the Atlantic east of Guyana and Trinidad.
The reason for this particular climatological behaviour lies in the weakening and northward shift of the north-east trade winds during the rainy season, often leading to extended periods of calm and shifting winds. Two important mechanisms concerning the conditions in the sea are affected by this process. The vertical exchange in the upper layers of the water column is greatly reduced so that the heat transfer downwards is practically inhibited. At the same time surface evaporation is similarly reduced and the surface temperature is then allowed to rise to a maximum. The other important process affected by the shift in the trade wind is the wind induced coastal upwelling which normally takes place along the Caribbean coast of Venezuela and Colombia. With the weakening or absence of the trade winds the upwelling ceases and the warmer surface water of the open Caribbean Sea finds its way back to the coast. Climatically this means that the normally prevailing stability of the air masses over the coastal region weakens or vanishes, thus allowing thermal convection to take place. Heavy rains may then follow. Air temperature and humidity remain high during this period.
2.1.1.3 Features of the oceanography of the Guianas, Trinidad, Venezuelan and Colombian coastal regions
The dominant feature of the oceanography of the adjacent seas of the mentioned coastal regions is the proximity of a major ocean current system: In the eastern areas the Guiana Current, which is an extention of the South Equatorial Current (SEC), in the northern areas (Venezuela and Colombia) the Carribbean Current, which in the coastal zone predominantly is an extention of the Guiana Current. Further north in the Caribbean Sea the North Equatorial Current has an increasing influence.
The mentioned ocean current system, flowing along a coast on its left side, indicates its presence by sloping density surfaces (isopycnals) upwards towards the coast in both the Atlantic and Caribbean regions. This feature is present at all times of the year, but is subject to regional and seasonal disturbances which will be related below.
Sloping isopycnals are normally taken as a sign of upwelling, although this may not necessarily be the case in the sense that the structure maintains a vertical velocity. If the structure is caused by local winds, upwelling in the true sense of the word is taking place. Otherwise, sloping isopycnals reaching the sea surface or near to it, may be looked at as a stationary feature, although responsible for the elevation of colder water, usually rich in nutrients, to higher levels in the sea. Combined with vertical eddy exchange the structure will complete the transport of nutrients to the euphotic zone. The effects on organic production will then be pronounced although not as intense as compared with actual winddriven upwelling. In the absence of windmixing organic production will normally be greatly reduced.
It will be evident from the following report that the mentioned processes play important but varying roles in determining the productivity of the costal zones in question.
As evident from the discussion above the oceanography of the survey area varies considerably along the mentioned coasts. A rough division may be made between the Atlantic and the Caribbean coasts due to 1) discharge of large amounts of freshwater to the Atlantic coasts of the Guianas (Suriname and Guyana) and 2) deflection of the incipient warm, low-salinity coastal water in the Caribbean to the north, making room for upwelling of a much colder (21°C) water along the Venezuelan coast. This difference in oceanographic characteristics reflects to some extent the behaviour of the major ocean currents of the region, but is more closely related to the difference in wind regimes affecting the coastal areas of the region. In the Atlantic the winds are predominantly blowing onto the coast, whereas in the Caribbean the winds are very much an alongshore phenomenon with only a slight on-shore component.
The paucity of precipitation in the Caribbean coastal zone makes this an area of high net evaporation, a process which hardly has much oceanographic significance in the open sea because of the rapid water exchange in the affected areas. Nearshore and inshore, however, this intense evaporation is of major significance to the environmental conditions of the coastal lagunes, many of them supporting large populations of domestically important fish and oyster resources. Drastic events happen occasionally when salinity exceeds marine life tolerance limits.
The exceptionally high primary production in the southeastern Caribbean area throughout most of the year is the cause of an intense “rain” of organic material to the deeper layers of the coastal waters. This leads to a significant load on the oxygen budget of the affected water masses. In some areas, notably the Cariaco Trench, the oxygen consumption exceeds the supply leading to anaerobic conditions in the deeper layers. The Cariaco Trench represents one of the few open ocean areas where the conditions allow large scale sedimentation of organic material.
2.1.2 Hydrography of the regions, as appearing from the “DR. FRIDTJOF NANSEN” scientific investigations 1988
2.1.2.1 The Guianas, Trinidad and Tobago
The observational material for this report is collected during four cruises during 1988: 21 January-14 February, 8-29 May, 11-28 August and 29 October-20 November. Standard oceanographic observations were carried out from a set of nearly fixed hydrographic sections which were repeated at the various cruises, thus facilitating comparison between the seasons.
The hydrographic observations are taken from 5 sections in the region. These are shown in Annex 1, DATA FILES, and are for convenience being referred to by the nearest well known coastal feature: Maroni, Demerara, Orinoco, Galeota and Tobago N (north).
2.1.2.2 Water masses of the region
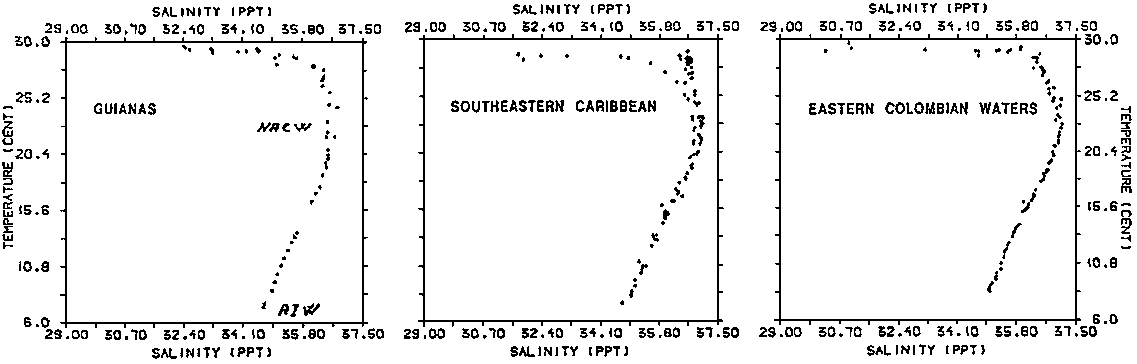
Observations from all sections are represented in the TS- diagram of Figure 2.1.1 showing values from stations off the continental shelf. This gives a picture of the source water masses of the region. We recognize the transition to the low salinity Antarctic Intermediate Water (AIW) in the deeper most part of the diagram. Above this we find the salinity maximum of the Central Water at 100 m depth, with salinities above 37 (most often observed only in the two southernmost sections). It is difficult to determine the origin of this water on purely hydrographic means, but the proximity of North Atlantic Central Water and certain features of the density structure indicate that this water actually is an intrusion of North Atlantic Central Water (NACW). The flow is obviously weak and strongly influenced by the ocean currents of the region, the Guiana Current, which is an extention of the South Atlantic Equatorial Current (SAEC), and the flow of SAIW at greater depths. The intrusion of NACW appears to be accentuated in a weak counter current off the shelf break. It is possible to understand this counter current energetically as a compensation current to erosion caused by vertical mixing with the water masses above and below
Above the NACW we find the Upper Water of the region, with highly varying characteristics in time and space. Typical features throughout all seasons are a lowering of salinity and increasing temperature towards the surface. The lowering of salinity may be tied in with influence of three different origins: 1) Amazon River water, which is responsible for an offshore salinity depression of vast extention, 2) equatorial excess precipitation and 3) local rivers, notably the Orinoco River. The influence of the local rivers is mostly quite distinct and limited in lateral extent, giving rise to bands of low-salinity water flowing along the coast.
2.1.2.3 Spatial distribution of salinity, temperature and oxygen concentrations
The spatial distributions of the variables above are presented in a series of vertical sections and in the form of horizontal charts of surface salinity and temperature (Annex 1, DATA FILES).
Salinity
Salinity is an outstanding parameter over the continental shelf and in the nearshore areas. The influence of the local rivers are particularly important in affecting the stability of the water column and thereby reducing the turbulent vertical exchange induced by wind energy at the surface. The presence of low salinity surface water along this coast must then be considered as affecting a reduced supply of nutrients, as otherwise would be provided by the moderate upwelling seen in most of the sections.
We have already pointed out the subsurface (100 m) salinity high found outside the shelf break. The isohalines are consistently sloping upwards onto and above the shelf, giving impression of upwelling on a large longitudinal (longshore) scale. This feature is present at all seasons and consistent with the existence of a major surface ocean current (Guiana Current). It is obvious, however, that the hydrostatic effect of large quantities of low salinity water along the coast will tend to amplify this feature and give impression of an enhanced upwelling. Although this is hardly the case in the common sense of the term, the elevation of water originating in the 100 m depth to levels up to 50 from the sea surface is a major process affecting primary production of the entire shelf region.
The lateral distribution of salinity is somewhat complex reflecting that a low-salinity northbound coastal current is dynamically unstable unless flowing with reduced velocity with respect to the underlying sea water. This particular dynamic condition is demonstrated clearly in the separation of the low-salinity water in local nearshore river extentions and an offshore regional band situated above and beyond the shelf break. The latter is a large scale feature which may well include an internal circulation with reduced or possibly reversed flow near the outer edge, whereas the inner edge may have a positive (northbound) velocity with respect to the underlying Guiana Current. The band as a whole may appear relatively passive.
It seems peculiar that the outer low-salinity band seen in the southernmost sections most of the year is weakening towards north and sometimes hardly discernible. This could be explained by a slight drift towards NE outside the area of observation. However, previous cruises in the area give indication of an intense vertical mixing which rapidly deteriorates the low-salinity band. This may suggest that the feature is rather of nearby origin, a conclusion which rules out the Amazon River as the origin of this water. In November this water is not seen in the sections, an indication that the flow may be passing outside the limit of observation, or that the source at this time of the year is particularly weak. This contrasts to the Orinoco River discharge which peaks in August and is still quite high in November.
The nearshore drift of low-salinity water is known to separate into two branches off the Serpents Mouth, one branch passing into the Gulf of Paria, the other forming a coastal flow on the windward side of Trinidad. The presented sections show that low-salinity water (mainly Orinoco) is present east of Trinidad at all times of the year, but greatly reduced in the winter and spring observations, reflecting the annual low in Orinoco River outflow in March.
Consistent with the present and previous observations, low-salinity water is found over large parts of the shelf area most of the year, but seemingly most predominant in the southernmost sections. Wind-induced upwelling does not seem to be important in this area, a conclusion which also is supported by previous investigations in this region. Vertical exchange is also expected to be greatly reduced by the shielding effect of the high stability of the upper layers. This observation is confirmed by the existence of generally high surface temperatures over the entire shelf area. It is possible, though, that the apparence of 26°C water over the shelf bottom (50 m) is sufficiently rich in nutrients to support a primary production in the lower parts of the water column.
Below the low-salinity bands of the surface layers, extending 10-30 m down, we should find South Atlantic Upper Water (SAUW), gradually changing into predominantly NACW indicated by salinities above 37‰. A peculiar feature of all sections is that the mentioned salinity maximum is found consistently 10-20 nm outside the shelf break. We interpret this feature as an indication of a somewhat stronger vertical mixing in the shelf break area, a condition which is commonly attributed to internal wave activity.
Temperature
The spatial distribution of temperature confirms by and large the observations and conclusions presented above. Shelf water temperatures are consistently high at all times of the year, but reveal the previously mentioned uplift at lower levels. This has been ascribed to the presence of the ocean currents of the region and possibly the hydrostatic effect of the low-salinity bands. Far offshore, normal temperatures of the region is about 29°C. The shelf water has a temperature which ranges from 25°C to 29°C of which the lowest values (25°C to 27°C) are found during the early parts of the year. The higher values are typical of the summer and fall observations. These are also consistent with a less pronounced sloping of the isotherms indicating a weaker “upwelling” of subsurface water. This ties naturally in with the behaviour of the Guiana Current which is known to be weakest at that time (Sep-Nov) of the year. At the same time the vertical mixing is reduced because of the generally weaker winds and larger content of fresh water in the upper parts of the water column.
Oxygen
The oxygen content of the shelf water is on the whole slightly lower but does not differ much from that of the adjacent ocean. The upper parts of the water column (0-50 m) have oxygen concentrations generally around 5 ml/l, whereas in the lower parts (50-100 m) concentrations vary from 4.5 to 3.5 ml/l, with a tendency towards the lower values (3.5 ml/l) to the north (Tobago). Occasionally the oxygen concentrations fall below 3.5 ml/l in the lower parts of the shelf water column, particularly in the northern sections. The behaviour is consistent with the discernible, but not too intense organic load of the region. Also the somewhat higher elevations of the entire water structure contributes to lower oxygen values encountered in the northern parts of the region. Outside the shelf break (100 m) oxygen concentrations diminish with depth towards a minimum of about 3 ml/l at the greatest measuring depth of 500 m. No particular oxygen feature can be recognized in connection with the salinity maximum (4 ml/l), nor with the counter current structure immediately below.
Seasonal variations are about 0.5 ml/l with the highest values (4.5-5.0 ml/l) of the shelf water found in the late fall and early spring observations.
2.1.3 Hydrography of the regions: Venezuela, north coast from Dragon's Mouths to Gulf of Venezuela
2.1.3.1 The hydrographic programme
Hydrographic observations are available from 4 cruises in this region: 15 February - 5 March, 30 May - 15 June, 29 August - 15 September and 21 November - 8 December. The cruise programme covered 6 hydrographic sections of which 5 were maintained in a NS direction. These are referred to by the nearest coastal feature: Pta. Caribe, Pta. del Taquien (Paria Peninsula), Puerto de la Cruz, Cabo Codera, Pta. Sabana and Gulf of Venezuela. The sixth section ran in a WNW direction from Paraguana Peninsula, joining the outer end of the section from Gulf of Venezuela. This programme was maintained with very little variation throughout the 1988 investigations with the exception of February when Cabo Codera and Pta. Sabana were omitted. In addition surface temperatures are available from thermograph recordings from the February and November cruises. These observations provide a continuous registration in sections of much closer spacing and are therefore very helpful in filling out the gaps existing in the regular hydrographic programme. Additional information on the oceanography of this coastal region is also provided by a section north of Guajira Peninsula, otherwise discussed in connection with the presentation of the westernmost (Colombian) region.
2.1.3.2 Water masses of the region
Representative data from the area have been plotted in the TS-diagrams of Figure 2.1.1. to show the characteristic water masses of the region. As could be expected the TS-configuration is not very different from those of the adjacent Atlantic Ocean, but it does show certain features typical of the Caribbean Sea and also the local influence of the Venezuelan shelf area.
2.1.3.3 Spatial distribution of salinity, temperature and oxygen concentration
The spatial distribution of salinity, temperature and oxygen concentrations is presented in a series of vertical sections as obtained from the regular hydrographic programme (Annex 1, DATA FILES).
Temperature
Sea surface temperatures are available in two charts from the first and the last cruises of the year (Annex 1, DATA FILES). Although only 3 months apart in the yearly cycle, these observations are almost extremes of the year. The February observations demonstrate intense upwelling with surface temperatures generally in the lower end of the range. At the beginning of December the same year the sea surface temperatures were close to the maximum of the year, although slightly lower. This latter difference is likely related to the winter reduction in solar radiation which at the end of November is significantly weaker than in August/September when it is at maximum. The distribution of surface temperatures does, however, indicate that upwelling is taking place also at this time of the year (Nov-Dec), but on the whole to a rather limited extent.
From the thermograph readings it is seen that spring time upwelling leads to a conspicuous distribution of temperature with coastal values down to the extreme low of 21°C. in local areas. So cold water is primarily present along the north coast of Paria Peninsula, but low temperatures are also found north and west of Araya Peninsula, and inside and west of Gulf of Cariaco.
Water of a temperature of 21°C is normally found in the adjacent Caribbean Sea at depths of about 150 m. Although most of the water reaching the sea surface stems from depths less than this, the upwelling represents a significant flux of nutrients to the euphotic zone, a fact which is demonstrated by the extreme turbidity and light green color of the coastal water.
Further west the spring time coastal water surface temperatures rise to a level of about 26°C with isolated maxima in Golfo Triste and Gulf of Venezuela (>27°C). Lower temperatures are found in the wake of Paraguana Peninsula, although much too high to indicate upwelling of significance. Thus, upwelling is not a characteristic feature of the surface water of the western part of this region.
In the late autumn the sea surface temperatures are generally higher. Two areas of lowered temperatures (< 25°C) can be recognised and must be taken as indicative of upwelling. The easternmost of these areas coincides with that of spring time extremes found along the northern coast of Paria Peninsula. The other is, surprisingly, a coastal strip east of La Guaira. The feature is likely connected to wakes caused by the particular topography of the area: the sharp bend of the coastline east of Cabo Codera and the shallow grounds south of and surrounding Tortuga Island. The temperature of both areas contrasts to far offshore values which exceed 27°C most of the year. Upwelling along the Venezuelan north coast is thus primarily a phenomenon of the Oriente.
The reason for the observed pattern of upwelling is to be found in two basic features: 1) Deflection of the Caribbean Current and 2) the behaviour of the wind regime in the Caribbean region. In contrast to the Guiana Current, which flows along the South American east coast, the extention in the Caribbean region, the Caribbean Current, is deflected towards north relative to the east-west direction of the coastline. For a while, the southern edge of the Caribbean Current seems to follow the continental shelf break causing the sloping isopycnals in very much the same fashion as for the Guiana Current in the Atlantic. However, with the flattening of the shelf break further west, the Caribbean Current is steadily loosing the identity of a coastal boundary current. Instead, it deflects towards north, leaving the shelf region of the northern coast of Venezuela steadily less influenced by its presence in the Caribbean Sea.
The characteristic sloping of the deep isopycnals (and isotherms) observed in this area (e.g. St. 91, Annex 1, DATA FILES) makes nutrient-rich and cold water available for (further) upwelling on the continental shelf. The latter process is directly related to a locally wind driven surface current which flows along the Venezuelan coast beginning at Dragon's Mouths. Deflection to the right by the rotation of the earth is compensated by a true vertical uplift of the lower part of the shelf water column. The two processes combine to give the conspicuous condition of upwelling along Paria Peninsula.
In spite of equally strong ENE winds further west, upwelling west of Margarita Island and Tortuga Island is invariably weaker in the process of bringing deep water to the surface. In the Puerto de la Cruz section (Annex 1, DATA FILES) the coldest water encountered is nearly 23°C. Further west the surface temperatures are steadily rising towards 27°C. typical of Gulf of Venezuela water. This failure to produce equally low surface temperatures can only be explained by the absence of the deep ocean current such as observed to follow the shelf break further east.
Salinity
Salinity is not a determining property of the mass field in the southern Caribbean except in the easternmost corner. Here, the well defined low-salinity water reveals its origin in the Orinoco River and other Guiana rivers, and contrasts to the widespread, but weakening influence of Amazon River in more northerly areas of south-eastern Caribbean Sea.
In the dry season, lasting from the beginning of the year through June, low-salinity surface water is present in the Caribbean in the form of a continuous band entering through Dragon's Mouths. Very little, if anything at all, shows up north of Trinidad. The low-salinity water from Gulf of Paria is known first to take a northerly (later in the year even NE) course, thereafter turning sharply towards NW, following or riding on the ocean current from the Tobago-Trinidad channel. We see this water in the Pta. Caribe section at St. 32 with salinities down to 33‰ and further west in May in the Pta. del Taquien section at Sts. 88 to 91, with a minimum of 35.6‰ at St. 90. The low salinity water is known to take a course north of Blanquilla Island indicating the general direction of the Caribbean Current which carries the low-salinity water far into the Caribbean Sea.
Later in the year, with the weakening of the trade winds and growing river outflow, low-salinity water enters the Caribbean both north of Trinidad and through Dragon's Mouths. The two bands seem to join downstream, but may maintain two salinity minima for some distance. The general direction of the flow seems very much the same as earlier in the year, but the extent and magnitude of the flow are clearly increased, particularly the part extending from the Trinidad-Tobago channel. In the Pta. Caribe section taken at the end of August an isolated low with possibly two minima was present at Sts. 167-168-169 with salinities below 30‰. Similar low salinities were present in the more westerly section Cabo Tres Puntas at the end of November, here with salinities below 30‰ at St. 255 and northwards. In both examples, the main salinity minimum is situated shortly outside (north of) the shelf break, whereas the inner minimum at all times is found immediately inside the shelf break.
The flow of low-salinity water in the southeast Caribbean is an exceptionally fine demonstration of the behaviour of the ocean currents of the region. The associated upwelling is demonstrated both in the form of presence of high salinities and low temperatures in the immediate vicinity of the coast.
It is also demonstrated that the absence of the Caribbean Current as a coastal current further west is the main reason for the weakening and finally disappearing upwelling, in spite of fairly strong longshore winds in that area.
The salinity maximum recognized in the Guyana sections in the Atlantic is also present in the south east Caribbean Sea. This maximum follows the general seasonal vertical fluctuations of the region, rising in the autumn to levels of about 120 m and descending to about 160 m in the spring, following the variations of the thickness of western equatorial Upper Water. In the coastal area local upwelling superimposes a variation which to a great extent counteracts the general trend of the region but with a slight shift in phase. High salinity water is then seen to rise towards the continental shelf and in places actually flood the shelf area. This was the case in the Oriente in May-June when moderate upwelling caused 37‰ water to invade the entire Puerto de la Cruz and Cariaco Trench area. This water is also recognized by its low temperature in the 21-24°C range.
It is possible that the extreme case of upwelling seen in February was the combined effect of the seasonal thinning of the Upper Water with a strong locally induced coastal upwelling. The salinity maximum was then at 75 m outside the shelf break and the water encountered over the shelf bottom was down to 18°C.
The structure of the density field along the north coast of eastern Venezuela may well imply a weak counter current running along the steep slope of the shoreline or off the shelf break. Such a flow is likely to be covered by a winddriven surface current and not easily discovered without proper instrumentation. The existence of a counter current is also indicated in the various sections further west as far as to Cabo Codera, possibly all the way to Pta. Sabana.
Oxygen concentration
The oxygen content of the water masses of the Caribbean Sea does not differ significantly from that of the adjacent Atlantic Ocean. Source water of the region is still the NA central Water occupying the upper 100 to 150 m of the water column and recognized by salinities above 37‰. At the surface the NACW is influenced by precipitation and low salinity coastal water.
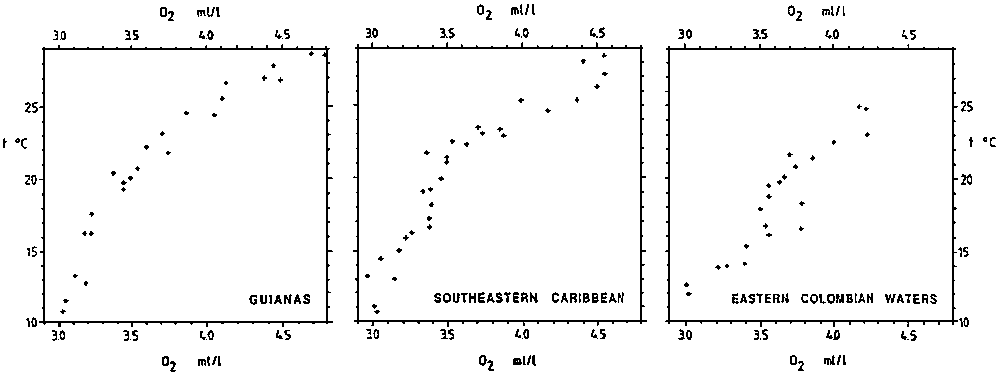
Oxygen values of the Upper Water of the Atlantic center around 4.5 ml/l at levels down to 100 m. Below, there is an increasing influence of the Antarctic Intermediate Water with oxygen concentrations down to below 3 ml/l. This picture also holds for the Caribbean area, but subject to a seasonal fluctuation containing also a signal of coastal influence. Far offshore values indicate source water characteristics such as presented in the T-O2 diagrams of Figure 2.1.2.
The most coherent T-O2 diagrams are those pertaining to the autumn cruises. Not only do we find fairly consistent T-O2 relationships, but there is also with very little variation between the different regions. The form of the T-O2 curves is nearly a straight line, a feature which can be interpreted as indicative of a uniform and relatively modest oxygen consumption in the water column. A slight curvature in the 18°C to 22°C range coincides with the main pycnocline and is likely related to enhanced organic decomposition in corresponding depth range. It is possible, though, that this water also is influenced by exchange with continental shelf water which is markedly reduced in oxygen content at similar depth range. In the Caribbean the curvature is developed into a weak minimum which is centered around the 20°C isotherm.
In the winter and spring the oxygen concentration in the lower range of the Central Water is much more variable than in the autumn, a feature which is interpreted as a result of springtime upwelling and enhanced organic production. Most of the production takes place over the continental shelf and leads to increased oxygen consumption in the lower range of the water column. A zone of oxygen minimum is developed, increasing westward from above 3.5 ml/l in the eastern Caribbean to less than 3.3 ml/l off Pta. Sabana, and also with a slight shift towards higher temperatures. Over the continental shelf oxygen concentrations are consistently lower in the lower part of the water column, occasionally becoming less than 3 ml/l. Similarly low values are also present over Cariaco Trench at all times, but with shifting depth levels. It is presumed that the Margarita Island-Araya Peninsula-Puerto de la Cruz area acts like a wake for the coastal flow and inhibits further aeration of the shelf water. Surface water rich in nutrients is, however, constantly being replenished to the upper layers, thereby maintaining high organic production.
Dynamical considerations
Certain features of the isotherms along the Venezuelan north coast indicate the existence of a weak countercurrent situated below the westward surface winddrift. The core seems to lie in the 50 to 100 m depth range and will thus appear as a bottom current over the continental shelf wherever this exists. In the Cariaco Trench area such a counter current may well support a general cyclonic circulation of deeper water.
2.1.4 Hydrography of the regions: Colombia from Guajira Peninsula to Sta. Marta
2.1.4.1 The hydrographic programme
The hydrographic programme consisted of stations taken at mostly three sections normal to the coast at locations off Guajira Peninsula, Riohacha and Rosario Islands at four cruises during 1988. In addition thermograph readings are available from the first and the fourth cruise, giving an extended picture of coastal water surface temperatures. Two hydrographic sections on the first cruise and one in the last cruise were omitted. The cruises took place 3-11 March, 14-21 June, 15-23 September and 6-11 December 1988.
2.1.4.2 Spatial distribution of salinity, temperature and oxygen concentration
The spatial distributions of the variables above are presented in a series of vertical sections and discussed in detail below.
Temperature
Spring time temperatures of the coastal water column are seen in the Guajira Peninsula section of Annex 1. DATA FILES. There is some indication of weak upwelling which may affect the inner coastal areas to some extent. When compared with the thermograph readings of Annex 1, DATA FILES the upwelling is seen to strengthen locally further west, but weakens again and disappears altogether in the western part of the survey area. Surface temperatures are down to less than 23°C. The local upwelling north and west of Guajira Peninsula must therefore be looked on as a local phenomenon, perhaps not even very persistent. The upwelling is likely related to a weak westward coastal current and amplified by the particular topography of the area.
The later cruises in the year confirm that the Guajira Peninsula area is a site of weak upwelling most of the year, but particularly weak in the summer and autumn observations. In December the upwelling strengthens and produces surface temperatures down to less than 24.5°C. This general pattern holds also for the areas further west but with steadily decreasing signs of upwelling, at times hardly discernible.
In the western part of the survey area, west of Cartagena, the surface temperatures are particularly high at all times of the year, reaching 29°C in the neighbourhood of Gulf of Morrosquillo.
Salinity
Salinity does not form characteristic features in this region. Far offshore a subsurface high above 37 ‰ is situated about 140 m depth in the late winter, but rising to about 100 m depth during the summer and weakening. The deepening follows the regional and equatorial winter convergence of Upper Water, as discussed in connection with Guyana waters. The slight salinity reduction in offshore waters towards the surface appears to be very much the same as in more easterly parts of the Caribbean and is apparently a general feature of the region. The upwelling discussed above tends to weaken this feature, lifting the salinity maximum towards the coast, but never much above the 100 m depth level.
Local influence on the salinity is not seen in this region except in the westernmost parts of the area where the salinity is down to below 32‰ (Rosario Islands). This water does not show up in more easterly sections, a fact which supports the idea of a westward coastal current along the Colombian coast. The few available sections from the Colombian coast do, however, suggest the existence of a weak eastward subsurface counter current, located in the 100-150 m depth range. The feature is present in all sections off Guajira Peninsula at all times, but generally so weak that it does not necessarily imply actual reversal of the velocity field. Even if an eastward coastal subsurface current were established as a more or less permanent phenomenon of the Caribbean coast of South America, it may well be limited to a thin band not reaching further out than 10 nm. The oceanographic implications of such a counter current are not clear.
Oxygen concentration
The oxygen concentrations offshore are very much in agreement with distributions further east. Outside the shelf break, concentrations vary from above 4.5 ml/l to below 3 ml/l at 300-400 m. In the early spring a weak minimum had developed in the 100-120 m depth range. The 3.5 ml/l surface was then above 100 m depth whereas it normally was found below 150 m, occasionally down to the 200 m depth level. It is possible that this reflects the plankton blooming in connection with wind induced spring time upwelling.
* This chapter is contributed by G. Bianchi, Institute of Fisheries Biology, University of Bergen.A study of the main demersal species associations in relation to environmental variables is being carried out using the trawl data of the R/V “DR. FRIDTJOF NANSEN”. The description given below is part of this work.
The analysis is mainly centered at the demersal assemblages. Pelagic species caught in the bottom trawl are also taken into account in their role as part of the trophic chain of the bottom-living species.
Species groups were analysed by a classification method, TWINSPAN (Two Way Indicator Species Analysis) and an ordination method, DECORANA (Detrended Correspondence Analysis).
2.2.1 Environmental factors affecting the shelf fauna
Environmental factors such as oceanographic conditions and type of bottom are of prime importance in controlling the distribution of shelf fauna.
A main feature of the northeast coast of South America, east of Trinidad, is the enormous water discharge from the Amazon, Essequibo and Orinoco rivers, carrying silts and soft mud which are partly deposited outside the river mouths and partly carried along the shelf by currents and tides. Apart from the typical brackish water environments associated to the river mouths, the freshwater output is so big as to influence large coastal areas. In the period of its maximum discharge, (June to December) the Orinoco is responsible for a zone of low salinity along the northeast Venezuelan coast and along the east coast of Trinidad. The effect of water discharge from rivers affects the shelf to about 40 to 60 m. The mud deposits are the substratum for a rich invertebrate fauna forming the main food item of many fish and several species of sharks and rays. At greater depths, sandy or rocky (old corals) grounds become predominant and the fauna changes accordingly to typical hard bottom lutjanid community. The shelf slopes gradually to about 100 m before it falls steeply into deeper waters. Probably for this reason, typical deep-shelf/upper slope groups are not found.
The thermocline does not play an important role in determining different faunistic regimes in these regions. The discontinuity layer is not sharp on this side of the Atlantic, its lower boundary quite deep. High temperatures (above 22°C) and high oxygen levels (above 3 ml/l) are found on the whole shelf throughout the year.
The Guiana Current is present throughout the year and causes an upward movement of water originating in the 100 m depth up to the 50 m level. Nutrients coming from this process and from the water discharge of the rivers affect the primary production of the entire shelf region. No wind-driven upwelling is present in this area as the main winds are actually blowing onshore.
The northern Venezuelan shelf, west of Trinidad is quite varied, both in bottom type and oceanographic conditions. Its eastern part is influenced by water from the Orinoco River coming through the Dragon's Mouths. The mud sediments found in the Carupano Depression originate from the plume of the Orinoco and are transported westward by the main current system. The Tortuga Bank, from Testigos Islands to Cabo Codera, is covered with sandy sediments and numerous areas of hard bottoms with typical hard bottom fish communities. Upwelling occurs along the north coast of Venezuela westward to approximately La Guaira, mainly in January-February. Signs of upwelling are also found west of the Guajira Peninsula, throughout the year but most conspicuous during the first part of the year.
Also here the thermocline is not sharp. Oxygen levels above 3 ml/l are found to at least 200 m depth throughout the area, except in the Cariaco Basin which is completely closed and anaerobic below 300 m. Oxygen levels of 1 ml/l were found at depths of about 150-200 m. No biological sampling was made in the deeper part of this basin.
The Gulf of Venezuela is protected from the open waters of the Caribbean by the Peninsula de Paraguana on the east and the Peninsula de Guajira on the west. The southern part of the Gulf is shallow (to about 30 m depth) and is connected to the Lake of Maracaibo, a brackish water environment receiving fresh water from the rivers and sea water entering from the mouth of the lake. Because of this connection and because of the bottom type, we find here species assemblages similar to those of the estuarine areas of the Atlantic coasts.
The Colombian shelf, from the Guajira Peninsula to the Gulf of Uraba, is quite varied, with alternating sandy, muddy and rocky bottoms. It is characterized by being quite shallow and abruptly falling to depths of one hundred to a few hundred metres. The shelf north of the Guajira Peninsula is very narrow, the bottom is sandy/muddy. Westward, from Cape Vela to just before Sta. Marta region the shelf widens again. Here the bottom is mainly sandy, with shell and coarse sand in some areas. Westward, and especially off Cartagena and south to the Gulf of Morrosquillo, there are frequent rocky/coral outcrops. In its west most section (Gulf of Uraba and adjacent areas) the bottom is quite muddy.
Weak upwelling occurs north and west of the Guajira Peninsula, more pronounced in winter and spring.
Because of the above characteristics the area will be analyzed according to its major and more distinctive ecological units: the region between the Maroni River to the Orinoco River, the northern coast of Venezuela (including the Tortuga Bank) and the Gulf of Venezuela. The Columbus Channel will be analysed under Trinidad and compared to the northern and eastern coasts of that island. The Colombian shelf includes several biotopes and for this reason will be discussed separately. The upper slope will also be treated in a separate section.
2.2.2 Species assemblages between the Maroni and the Orinoco rivers
The very shallow waters are usually not very well sampled by the R/V “DR. FRIDTJOF NANSEN” because of operational limitations. However, several stations between 15 and 20 m were carried out in this region. Species typical of these waters and especially associated with waters with lower salinity were the stardrums Stellifer sp., the shortfin corvina Isopisthus parvipinnis, the king weakfish Macrodon ancylodon, the spicule anchovy, Anchoa spinifer and, in smaller quantities, two species of anchovies of the genus Anchoviella, the Guiana longfin herring Odontognathus mucronatus, the dogtooth herring Chirocentrodon bleekerianus, the green weakfish Cynoscion virescens, and three species of catfishes Arius grandicassis, Bagre marinus and B. bagre. Batoid fishes are also common inhabitants of shallow muddy bottoms and brackish waters. Among these, the smooth butterfly ray Gymnura microura and the longnose stingray Dasyatis guttata appeared to be common inhabitants of the shallow inshore waters. All the species mentioned above feed on small fishes, shrimps and other bottom invertebrates which are particularly abundant in the estuarine systems and muddy bottoms. Several species of shrimps are abundant here some of which are restricted to the very shallow waters: the Atlantic seabob, Xiphopenaeus kroyeri, the southern brown shrimp Penaeus subtilis and the southern white shrimp Penaeus schmitti. The redspotted shrimp Penaeus brasiliensis and the southern pink shrimp P. notialis were found in deeper waters. The different behaviour shown by the two groups (higher catch rates during daytime for the former while the opposite is true for the latter ones) is probably related to different conditions of water turbidity in their respective areas of distribution.
Other species with a wider depth distribution were often associated with the above species: the whitemouth croaker Micropogonias furnieri, the American pellona, Pellona harroweri and large predators like the hairtail Trichiurus lepturus, the Caribbean sharp-nose shark Rhizoprionodon porosus and the sierra Spanish mackerel Scomberomorus brasiliensis. The round sardinella Sardinella aurita was mainly caught by bottom trawl on the deeper part of the shelf (60 to 90 m). This species is known to prefer clear saline waters with temperatures below 24°C. Off the Guyanas shelf these conditions are indeed found only along the outer shelf, near to its edge. Small sardinella (average length between 4 and 7 cm), was found at depths of 25-40 m in the course of the fourth survey in November. This might indicate a spawning season in June-July.
In the intermediate shelf area the lane snapper Lutjanus synagris the slender inshore squid Loligo plei, the inshore lizardfish Synodus foetens, the bolo seabass Diplectrum radiale and the bluntnose jack Hemicaranx amblyrhynchus were commonly found together with pelagic or semipelagic larger predators such as the king mackerel Scomberomorus cavalla and the guachanche barracuda Sphyraena guachancho. Smaller pelagics were also characteristic of this zone: the Atlantic bumper Chloroscombrus chrysurus and the Atlantic threadherring Opisthonema oglinum.
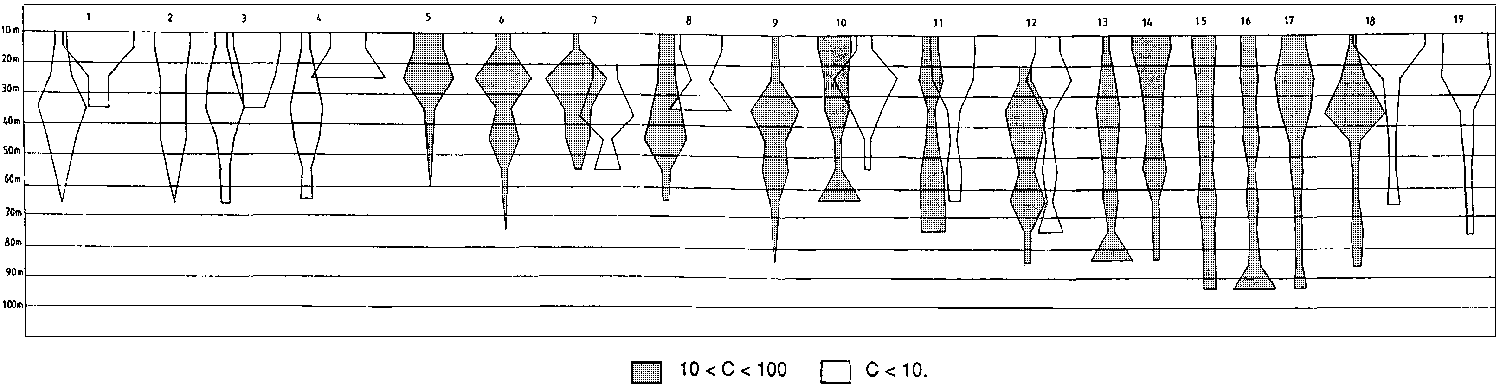
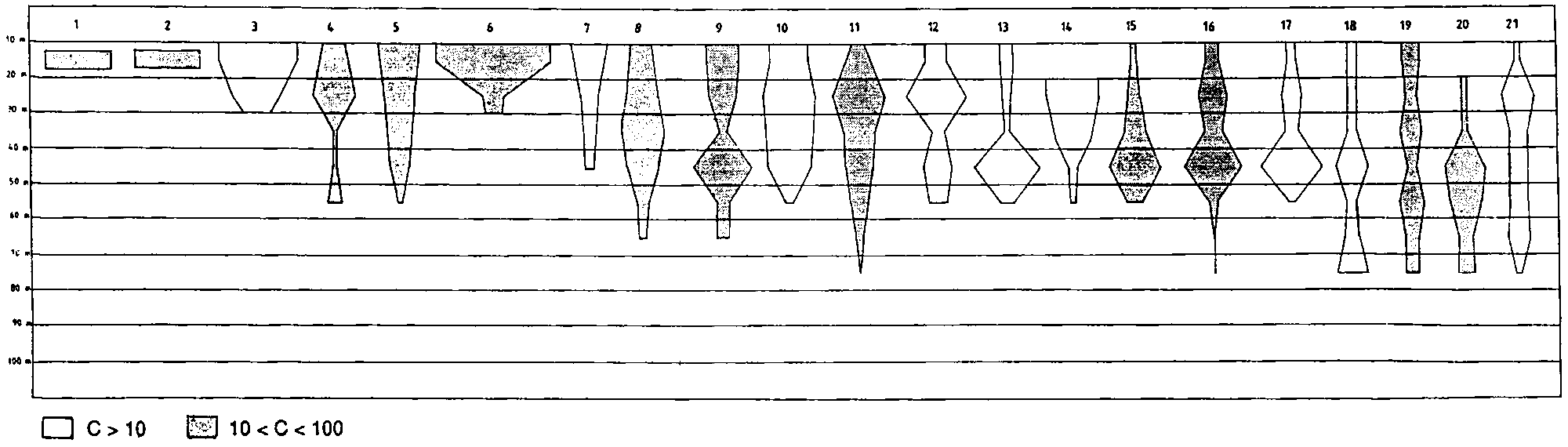
Figure 2.2.1 shows the bathymetric distribution of the above species.
In deeper waters to the edge of the continental shelf species assemblages change considerably, especially in the deeper shelf off Suriname and Guyana. Here the catches consisted mainly of “red fish”. Dominant in this region are the vermillion snapper Rhomboplites aurorubens, the cardinal snapper Pristipomoides macrophthalmus and the southern red snapper Lutjanus purpureus. The Atlantic bigeye Priacanthus arenatus is also associated with this group. This species, reported in the literature as living around reefs, solitary and forming small aggregations in the 15-75 m bottom range, is one of the dominant species in the deeper part of the shelf, on hard bottoms and may be caught by trawl in large quantities. This has been observed also in the eastern Atlantic, especially off Congo and Gabon. The dwarf goatfish, Upeneus parvus was also mainly associated with this group (see Figure 2.2.2).
An additional outline for the same species is given in the case of difference in depth distribution in the warm season. Different shades indicate different average catch rates (in kg/h):Differences in the bathymetric distribution of several species of the intermediate waters seemed to occur especially in August (third survey). This is the only time of the year when the trade winds stop blowing, shallow waters become clearer and there seems to be a general tendency for the above species to move shoreward.1) Bagre bagre 2) Stellifer sp. 3) Isopisthus parvipinnis 4) Macrodon ancylodon 5) Anchoa spinifer 6) Xiphopenaeus kroyeri 7) Lutjanus synagris 8) Cynoscion virescens 9) C. jamaicensis 10) Pellona harroweri 11) Hemicaranx amblyrhynchus 12) Selene setapinnis 13) Chloroscombrus chrysurus 14) Opisthonema oglinum 15) Scomberomorus cavalla 16) Peprilus paru 17) Trichiurus lepturus 18) Sphyraena guachancho 19) Loligo plei
Northwards, the deeper shelf outside the Orinoco delta, seemed to support a different fauna. Although some species of the “red fish” group were present they were not dominant and other species more typical of muddy bottoms appeared. The toadfish Porichthys pauciradiatus, the southern pink shrimp Penaeus notialis, the Mexican flounder Cyclopsetta chittendeni the seabass Serranus atrobranchus seemed to be typical of this area.
2.2.3 Species assemblages around Trinidad
The analysis is mainly based on survey 4 (13-20 November) because of the higher survey intensity and a greater coverage which allows comparison between the sides of the island: the north and east coasts and Columbus Channel. The Gulf of Paria, with its special shallow water system, is excluded from the analysis because no stations were carried out there. Survey 2 (23-27 May), the second richest survey in stations, was also used to grossly compare the two different main seasons. A detailed analysis of seasonal variations in species assemblages was, however, not possible because of the differences in number and position of the stations in the two surveys.
Three main groups of species can be identified: 1) those occurring in shallow waters in connection with river estuaries (a) those found along the north and northeast coast, b) those found along the Venezuelan coast); 2) those species typical of soft bottoms, in slightly deeper waters (a) of Columbus Channel b) along the northern coast); 3) “red” species of the deeper shelf, mainly on hard bottoms (a) of the northern shelf and b) of the eastern shelf. A total of about 250 species were caught on the shelf. Table 2.2.1 gives the list of the main species caught on the Trinidad shelf and their presence in each group is indicated with an asterisk.
Fig. 2.2.2 Depth distribution of main deeper shelf species - Maroni to Orinoco River.
An additional outline for the same species is given in the case of difference in depth distribution in the warm season. Different shades indicate different average catch rates (in kg/h):Table 2.2.1 Occurrence of the main species caught on the Trinidad shelf - G1, G2 and G3 indicate the main groups. For further explanations see Text.1) Rhomboplites aurorubens 2) Pristipomoides macrophthalmus 3) Lutjanus purpureus 4) Lutjanus analis 5) Upeneus parvus 6) Priacanthus arenatus 7) Sardinella aurita.
|
|
G1 |
G2 |
G3 |
|||
|
a |
b |
a |
b |
a |
b |
|
|
Penaeus subtilis |
* |
* |
* |
* |
|
|
|
Isopisthus parvipinnis |
* |
* |
|
|
|
|
|
Larimus breviceps |
* |
|
|
|
|
|
|
Stellifer spp. |
* |
* |
|
|
|
|
|
Xiphopenaeus kroyeri |
* |
* |
|
|
|
|
|
Macrodon ancylodon |
* |
* |
|
|
|
|
|
Anchoa spinifer |
|
* |
|
|
|
|
|
Umbrina coroides |
* |
|
|
|
|
|
|
Gymnura microura |
* |
|
|
|
|
|
|
Odonthognathus mucronatus |
* |
|
|
|
|
|
|
Polydactylus virginicus |
* |
|
|
|
|
|
|
Bagre marinus |
* |
* |
* |
* |
|
|
|
Pellona harroweri |
* |
* |
* |
|
|
|
|
Cynoscion steindachneri |
* |
* |
* |
|
|
|
|
Anchoviella brevirostris |
|
* |
* |
|
|
|
|
Trachypenaeus spp. |
|
* |
* |
* |
|
|
|
Lepophidium profundorum |
|
* |
* |
* |
|
|
|
Anchoviel. lepidentostole |
* |
* |
* |
|
|
|
|
Anchoa spp. |
|
|
* |
|
|
|
|
Eucinostomus spp. |
|
|
* |
* |
* |
* |
|
Porichthys pauciradiatus |
|
* |
* |
* |
|
|
|
Penaeus brasiliensis |
|
* |
* |
* |
|
|
|
Penaeus notialis |
|
* |
* |
* |
|
|
|
Ctenoscia. gracilicirrhus |
|
* |
* |
* |
* |
|
|
Cynoscion jamaicensis |
|
* |
* |
* |
* |
|
|
Lutjanus synagris |
* |
* |
* |
* |
* |
* |
|
Chirocentr. bleekerianus |
|
* |
* |
* |
* |
|
|
Synodus foetens |
|
|
* |
* |
|
* |
|
Prionotus punctatus |
* |
* |
* |
* |
* |
|
|
Trichiurus lepturus |
* |
* |
* |
* |
* |
* |
|
Micropogonias furnieri |
* |
* |
* |
* |
* |
|
|
Harengula jaguana |
* |
* |
* |
* |
|
* |
|
Pomatomus saltatrix |
|
* |
* |
* |
* |
* |
|
Mustelus higmani |
* |
* |
* |
* |
* |
* |
|
Peprilus paru |
* |
* |
* |
* |
* |
* |
|
Opisthonema oglinum |
* |
* |
* |
* |
* |
* |
|
Loligo pealei |
|
* |
* |
* |
* |
* |
|
Selene setapinnis |
* |
* |
* |
* |
* |
* |
|
Lutjanus purpureus |
|
|
* |
* |
* |
* |
|
Rhizoprionodon porosus |
* |
* |
* |
* |
* |
* |
|
Chloroscombrus orqueta |
* |
|
* |
* |
* |
* |
|
Scomberomorus cavalla |
|
|
* |
|
* |
* |
|
Loligo plei |
|
|
|
|
* |
* |
|
Sphyraena guachancho |
|
|
* |
|
* |
* |
|
Sphyraena picudilla |
|
|
|
|
* |
* |
|
Selar crumenophthalmus |
|
|
|
|
* |
|
|
Upeneus parvus |
|
|
|
|
* |
* |
|
Trachurus lathami |
|
|
|
* |
* |
* |
|
Pristipom. macrophthalmus |
|
|
|
|
* |
* |
|
Priacanthus arenatus |
|
|
|
|
* |
* |
|
Rhomboplites aurorubens |
|
|
|
|
* |
* |
|
Decapterus punctatus |
|
|
|
|
|
* |
|
Calamus bajonado |
|
|
|
|
|
* |
|
Lutjanus analis |
|
|
|
|
|
* |
|
Alutera monoceros |
|
|
|
|
|
* |
|
Pomacanthus paru |
|
|
|
|
|
* |
The slightly deeper part of the Columbus Channel, the southern coast of Trinidad and some stations of the north at similar depths, are characterized by the following species: Cynoscion jamaicensis, Ctenosciaena gracilicirrhus, Eucinostomus spp. and Lutjanus synagris, Penaeus brasiliensis and P. notialis.
Species common to the first two groups were: Bagre marinus, Pellona harroweri, Harengula jaguana, Cynoscion virescens, Anchoviella brevirostris and A. lepidentostole. In the catches of the night stations were also caught Porichthys nautopedium, Lepophidium profundorum, Penaeus notialis and Trachypenaeus sp.
The stations more offshore, down to depths of about 90 m, differed for holding species of more typically hard bottoms. Apart from the pelagic species (Scomberomorus cavalla, Sphyraena picudilla and S. guachancho) Upeneus parvus, Pristipomoides macrophthalmus and Rhomboplites aurorubens were the species typical of these zones. Priacanthus arenatus was also associated but this species was found only in the north. A few stations off the southeastern coast of Trinidad, at around 30 m depth, showed a community typical of hard bottoms also associated with corals. Species like Calamus bajonado, Lutjanus analis, Alutera monoceros and Pomacanthus paru were caught here. The pelagic Trachurus lathami and Decapterus punctatus were also caught in the bottom trawl in the more offshore waters, the former mainly in the north, the latter in the east, in both surveys.
Ubiquitous species, present in all groups were Chirocentrodon bleekerianus, Trichiurus lepturus, Peprilus paru, Opisthonema oglinum, Chloroscombrus chrysurus, Mustelus higmani and Micropogonias furnieri.
As already mentioned, a detailed comparison between the two surveys (survey 2 and 4) is not possible because of a different coverage and survey intensity of the two surveys. It seems that some of those species found in connection with estuaries and river mouths, were more abundant and widespread in the course of survey 4, at the end of the rainy season when the influence of the water coming from the rivers is at its maximum. Isopisthus parvipinnis, Stellifer sp., Penaeus subtilis, Xiphopenaeus kroyeri were indeed most common and abundant at this time of the year.
All the deep-water stations will be treated together under the special section “the slope”.
2.2.4 Species assemblages of the northern coast of Venezuela, from Dragon's Mouths to Cabo Codera (excluding Tortuga Bank)
The eastern part of the northern Venezuelan coast is strongly influenced by upwelling almost throughout the year as a consequence of the deflection to the north of the Caribbean Current and of local winds blowing parallel to the coast. This makes this region the most productive of the Venezuelan coast and this is reflected in the large concentrations of small pelagics, mainly Sardinella aurita, found here.
The demersal fauna of the shallow coastal waters of the northern coast of the Araya Peninsula consists mainly of species with preference for muddy bottoms and associated to brackish water environments: the gafftopsail sea catfish Bagre marinus, the barbel drum Ctenosciaena gracilicirrhus, the whitemouth croaker Micropogonias furnieri and the corocoro grunt Orthopristis ruber. Since no important rivers are found in this area, environmental conditions favourable to this fauna might actually be generated by water originating in the Gulf of Paria and transported westward by the coastal current originating at Dragon's Mouths. Small pelagics associated with the above species in the bottom trawl were the Atlantic pellona, the scaled herring and the dogtooth herring.
Among the predators were the guachanche barracuda, hairtails and the smalltail shark Carcharhinus porosus.
2.2.5 Tortuga Bank and the west coast to the eastern coast of the Paraguana Peninsula
On the sand and hard bottoms of the Tortuga Bank and all along the west coast, at intermediate depths widespread and occasionally dominant are the dwarf goatfish Upeneus parvus, the Atlantic bigeye Priacanthus arenatus, the cardinal snapper Pristipomoides macrophthalmus, the vermilion snapper Rhomboplites aurorubens and the bronzestripe grunt Haemulon boschmae together with other semipelagic species like the slender inshore squid Loligo plei, the rough scad Trachurus lathami, the chub mackerel Scomber japonicus and the longfin inshore squid Loligo pealei.
The shelf in proximity of the coast differs, however, from the Tortuga Bank for holding, besides the widespread species, also some which are rare or absent in the latter. Among these the silver mojarra Eucinostomus argenteus, the southern red snapper, Lutjanus purpureus, the seabass Diplectrum bivittatum, the mutton snapper Lutjanus analis, the pluma porgy Calamus pennatula and the lane snapper L. synagris. Nursery areas for the young of the above species are usually shallow coastal areas, which could explain their absence on the offshore submarine bank. The bigeye scad Selar crumenophthalmus, which is also an inshore shallow water species, was also characteristic of the coastal shelf.
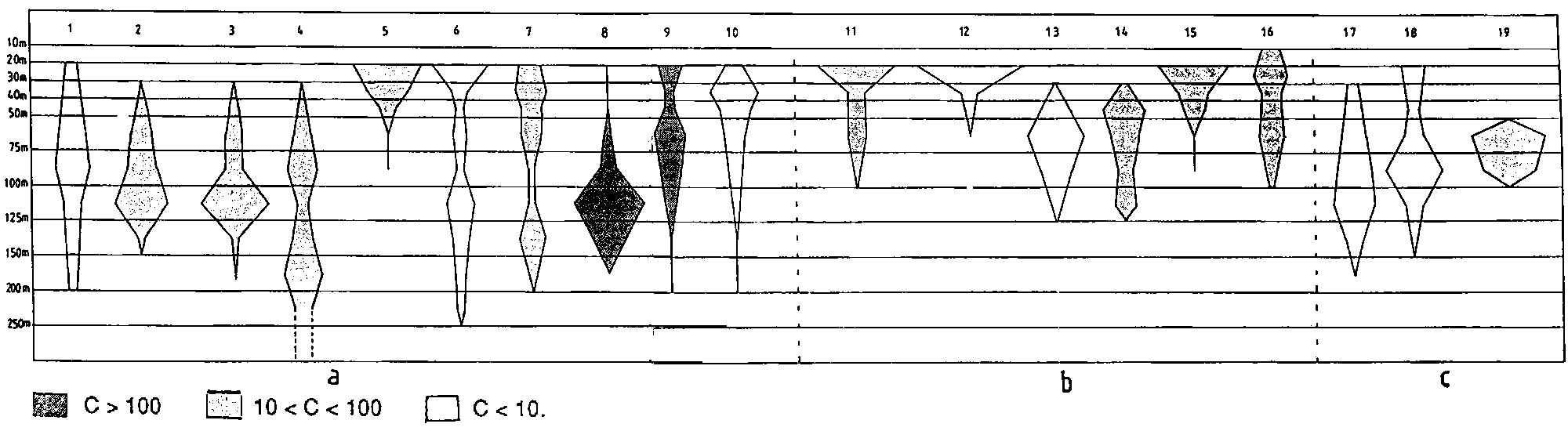
Figure 2.2.3 shows the bathymetric distribution of the above species.
a) Most ubiquitous species b) Species more typical of coastal areas c) Species more typical of the bank. Different shades indicate different average catch rates (in kg/h):a - 1) Upeneus parvus 2) Priacanthus arenatus 3) Rhomboplites aurorubens 4) Pristipomoides macrophthalmus 5) Haemulon boschmae 6) Loligo pealei 7) L. plei 8) Trachurus lathami 9) Sardinella aurita 10) Dactylopterus volitansb - 11) Eucinostomus argenteus 12) Diplectrum bivittatum 13) Lutjanus purpureus 14) Lutjanus analis 15) Calamus pennatula 16) Lutjanus synagris
c - 17) Synodus intermedius 18) Trachinocephalus myops 19) Haemulon striatum
2.2.6 Gulf of Venezuela
The fauna of this area greatly resembles that found in the shallow waters in proximity of the large estuarine systems of the Atlantic coast, the connection to Lake Maracaibo and the presence of soft grounds being the factors responsible for this. In the shallower part of the Gulf are the Atlantic bumper, hairtails, the Atlantic thread herring, dogtooth herring, spicule anchovy, the American pellona and the American harvestfish and the thumbstall squid Lolliguncula brevis. The purely demersal fauna seems to be less rich in number of species and consisted mainly of the shortfin corvina and the whitemouth croaker. This area has constituted since the '50ies important fishing grounds for the shrimp fishery. The white shrimp (Penaeus schmitti) and the Atlantic seabob seemed to be restricted to the very shallow waters in the south west part of the Gulf. The pink shrimp, Penaeus notialis and the redspotted shrimp Penaeus brasiliensis occurred in the deeper part of the Gulf together with other species characteristic of sandy-mud bottoms of intermediate waters like the cusk eel Lepophidium profundorum, the conger eels of the genus Hildebrandia and the corocoro grunt Orthopristis ruber.
Figure 2.2.4 shows the bathymetric distribution of the main species found in this area.
2.2.7 Colombia
The depth gradient is usually the most meaningful in explaining major differences in species composition of the marine fauna, at least on the continental shelf and slope areas. In the case of Colombia, however, because of the nature of the shelf, i.e. relatively shallow with an abrupt fall to greater depths, the stations do not cover a wide depth range and major differences in species composition can instead be related to other factors such as type of bottom and occurrence of upwelling which, although weak, does occur north and west of the Guajira Peninsula.
The stations carried out north of the Guajira Peninsula had an average depth of about 50 m. Demersal species commonly caught here were Lutjanus synagris and L. analis, Rhomboplites aurorubens, Pristipomoides macrophthalmus, Haemulon aureolineatum, Loligo plei and L. pealei. Demersal predators like Carcharhinus porosus and Sphyrna lewini were also occasionally caught. Several pelagic species also entered the bottom trawl, during daytime, especially in the first three surveys while they were much less conspicuously represented in the fourth survey: Sardinella aurita, Trachurus lathami, Scomber japonicus and Sphyraena picudilla.
West of the Guajira Peninsula the shelf widens. The bottom is mainly sandy, mixed with shells or coarse sand. The species caught here include a group typically associated to hard bottoms (Calamus pennatula, Balistes capriscus, Lactophrys trigonus, Acanthostracion spp., Pomacanthus paru, Lutjanus analis, Upeneus parvus, Rhomboplites aurorubens and Priacanthus arenatus, the first five species found mainly in shallow waters of about 20-25 m); a group of more ubiquitous species (Lutjanus synagris, Haemulon aureolineatum and Eucinostomus argenteus) and several pelagic species caught in the bottom trawl at day time, usually in the deeper part of the shelf (Sardinella aurita and Decapterus macarellus). The presence of these pelagic species north and west of the Guajira Peninsula is clearly related to the weak upwelling present almost throughout the year in this area.
The area from about Sta. Marta to the Gulf of Morrosquillo holds a type of fauna quite similar to the one described above but with the main difference that small pelagics were missing.
A more dramatic change occurs south of the Gulf of Morrosquillo, where we find, at depths of about 10-50 m, the typical fauna of soft bottoms and of areas influenced by the water discharge of large rivers: Chirocentrodon bleekerianus, Bagre marinus, Lycengraulis poeyi, Micropogonias furnieri, Selene setapinnis, Lolliguncula brevis, Penaeus notialis, Chloroscombrus chrysurus, Opisthonema oglinum and Trichiurus lepturus. Lutjanus synagris and Eucinostomus argenteus were also present.
Fig. 2.2.4 Depth distribution of main species in the Gulf of Venezuela.
Different shades indicate different average catch rates (in kg/h):1) Isopisthus altipinnis 2) Xiphopenaeus kroyeri 3) Penaeus schmitti 4) Pellona harroweri 5) Micropogonias furnieri 6) Anchoa spinifer 7) Harengula jaguana 8) Opisthonema oglinum 9) Chirocentrodon bleekerianus 10) Anchoviella lepidentostole 11) Chloroscombrus chrysurus 12) Scomberomorus brasiliensis 13) Selene setapinnis 14) Orthopristis ruber 15) Peprilus paru 16) Trichiurus lepturus 17) Lolliguncula brevis 18) Penaeus notialis 19) Lutjanus synagris 20) Sphyraena picudilla 21) Eucinostomus argenteus.
2.2.8 The slope
A total of 107 bottom trawl stations were carried out on the upper slope in the course of the four surveys, ranging from 200 to almost 800 m, mainly off Suriname, the southeast coast of Trinidad and outside the Orinoco, and off northern Venezuela and Colombia. About two thirds of the stations were carried out during nightime, when many deepwater species swim toward the surface. For this reason the following analysis might suffer of an important sampling limitation.
Deepwater stations from all surveys were analyzed simultaneously to obtain a better coverage of the area. Also, no important seasonal changes should occur at those depths. Correlation with depth and geographical position of the stations (longitude) showed that major changes occur mainly with depth while no real zoogeographic differences were detected.
Faunal changes along an environmental gradient are seldom sharp. Usually species gradually disappear while new species appear. However, although the above was true for most species, a significant change seems to occur at about 400 m depth were a break in the distribution of common species occurs. Polymixia lowei, Pontinus longispinis and Argentina sp. have at that depth their lower boundary while Benthodesmus thenuis, Plesiopenaeus edwardsianus and the isopod Bathynomus giganteus were usually found at depths greater than 400 m. The latter species is the largest living isopod. It is reported as pelagic, but it was caught only in bottom trawls, both during daytime and nighttime, in the course of these surveys.
A few species were ubiquitous and with a very wide depth range, found from the edge of the continental shelf to the deepest of the surveyed waters: Steindachneria argentea, Synagrops spp., Peristedion spp., representatives of the family Percophidae.
The above two major groups of stations/species, occurring above and below 400 m can be further divided.
Three well defined groups can be described within the stations below 400 m: a group of 13 stations off Venezuela, with an average depth of 441 m (SD: 42) with three species found consistently and almost exclusively there: Physiculus fulvus, Bathyclupea sp. and Aristeomorpha foliacea. In common with deeper stations this group has Bathynomus giganteus and Benthodesmus tenuis while in common with shallower stations were Peristedion spp., Steindachneria argentea, Illex sp. and Percophidae. Synagrops was practically not found here. A total of 91 species were identified from these stations.
Another group consists of stations from the Orinoco region, with an average depth of 603 m. (SD: 64), but also including a few stations from Suriname and Venezuela. Dominant in the catches was Synagrops sp., together with Bathynomus giganteus, Plesiopenaeus edwardsianus. Other species were Glyphocrangon sp., Nematocarcinus rotundus and Benthodesmus tenuis.
The 10 deepest stations, with average depth of 752 m (SD: 50) are characterized by two macrourid (grenadier) species only found here: Nezumia cyrano and Coelorhinchus occa. Other common species are those found below 400 m.
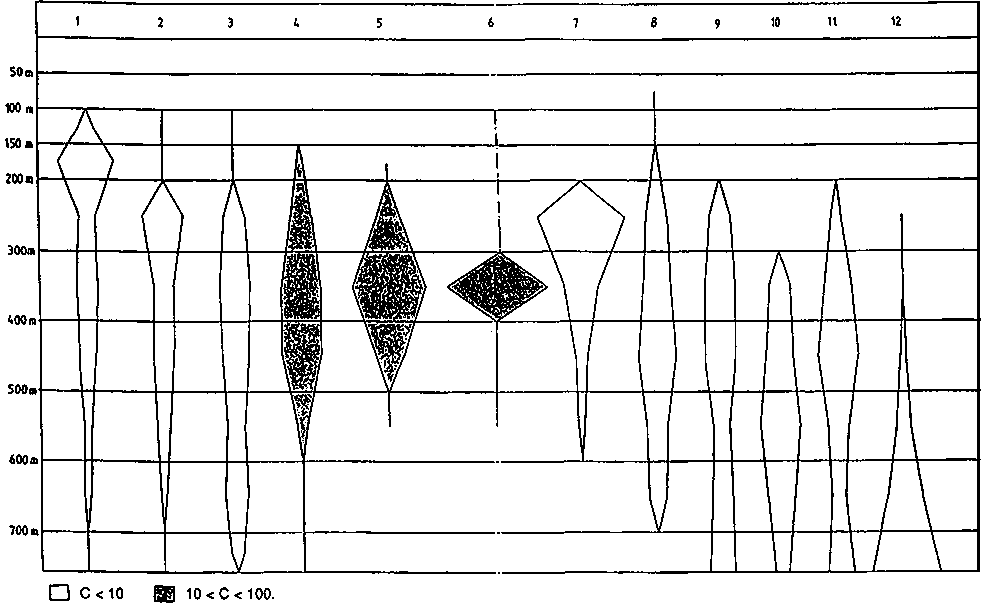
Fig. 2.2.5 Depth distribution of main slope species.
Different shades indicate different average catch rates (in kg/h):1) Peristedion sp. 2) Illex sp. 3) Synagrops sp. 4) Steindachneria argentea 5) Polymixia lowei 6) Ariomma bondi 7) Penaeopsis serrata 8) Solenocera sp. 9) Macrouridae 10) Bathynomus giganteus 11) Pleoticus robustus 12) Plesiopenaeus edwardsianus.
Also for the stations of the group between 200 and 400 m, depth is the main gradient along which species composition changes. The shallower stations (average depth of 250 m) hold several species also found on the shelf such as Lepophidium profundorum, Pristipomoides macrophthalmus, Antigonia capros and Trachurus lathami.
In tropical and subtropical regions most of the commercial types of small pelagic fish and all commercial demersal fish are found within the continental shelf areas of the sea. The extent and configuration of the shelf areas is thus important for the existence and distribution of these categories of resources.
Table 2.3.1 shows approximate estimates of the length of the shelf and the areal extentions of the continental shelf within depth ranges for each of the countries in the region. Special estimates are made for sub-regions where the resources will be analysed separately in the following. These include the Venezuelan shelf off the Orinoco estuary, the Joint Fishing Zone south of Trinidad and north of Venezuela, Venezuela Oriente to Cabo Codera, Venezuela West to Peninsula de Paraguana and the Gulf of Venezuela. Venezuela has by far the most extensive shelf area with more than 40 000 nm2, Suriname follows with about 19 000 nm2, Guyana has about 17 000 nm2 and Colombia about 12 000 nm2.
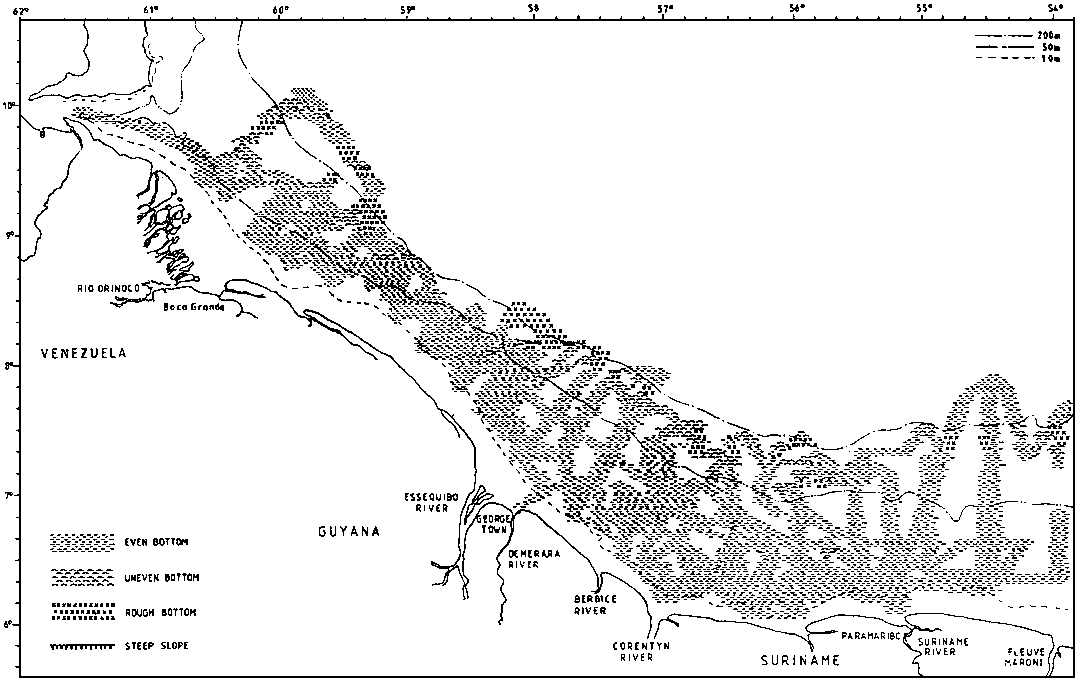
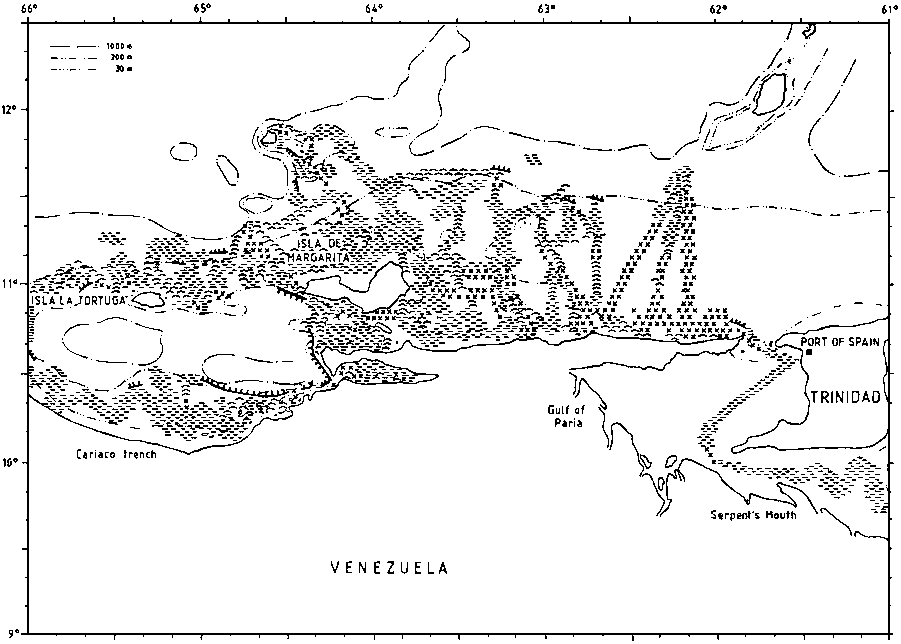
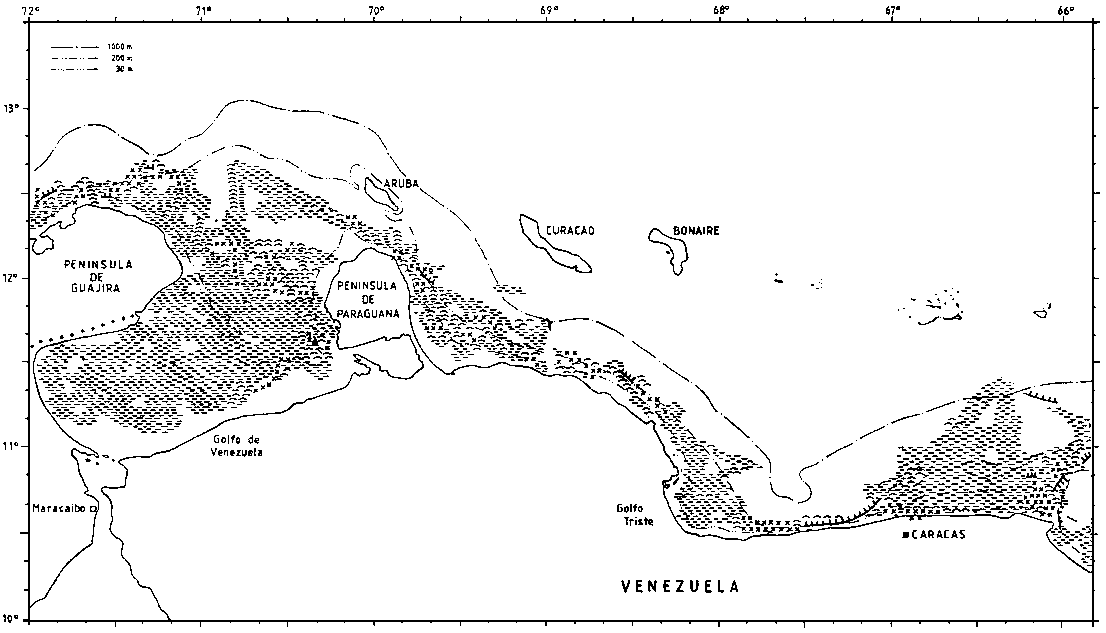
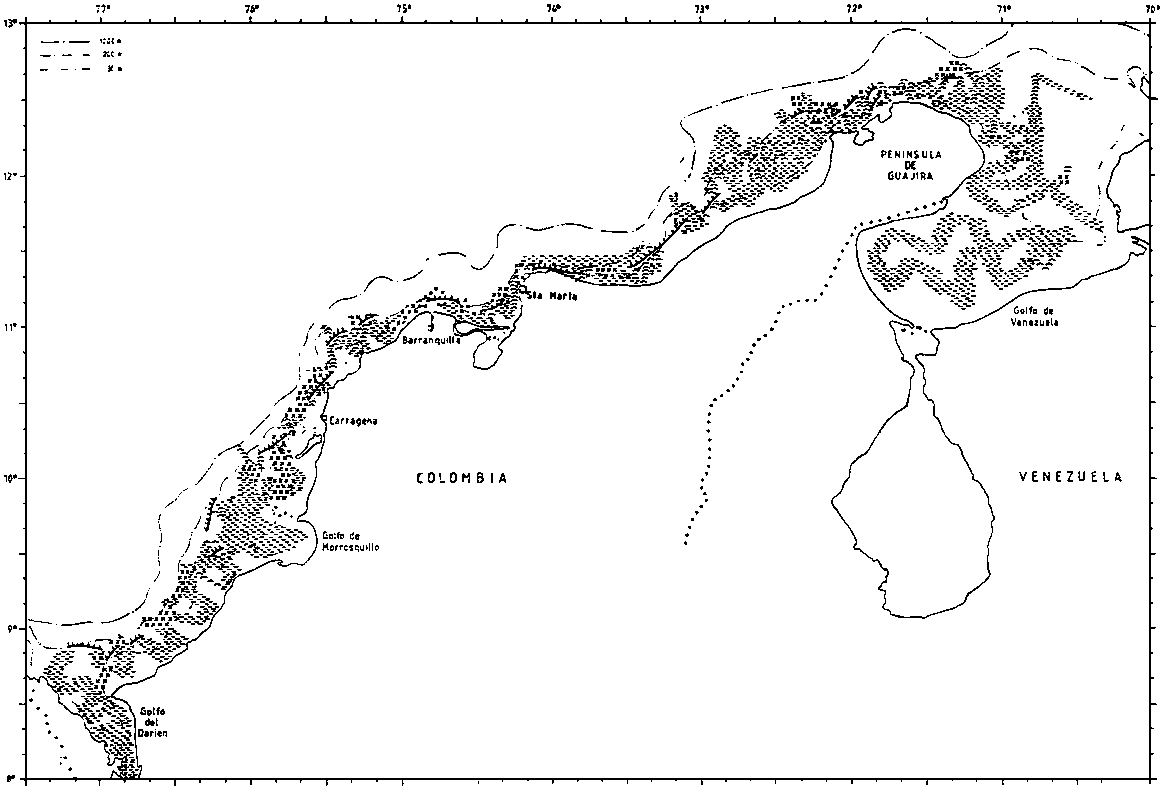
The type of bottom is of interest for fisheries and bottom character was observed along the survey tracks on the basis of an examination of the echo sounder diagrams. The resolution of these observations is 5 nm and since there is also some navigational uncertainty the precision of the charts produced will be limited. Four categories were used: smooth even bottom, relatively smooth but uneven bottom, rough bottom and very steep bottom. The last two are considered unsuitable for bottom trawling. But also the first category, even smooth bottom may be unsuitable for trawling e.g. on muddy very soft bottom.
Figure 2.3.1 shows the data on bottom character deriving from plots along the course lines and some interpolation. The shelf off the Guyanas is 40-90 nm wide and gently sloping out to about 100 m. There is then a relatively steep slope, but often with reduced gradient from 300-400 m and deeper. Most of the bottom is smooth and even, but there is a belt of sharp rough bottom at about 100 m inside the slope edge. Soft bottom can present problems for heavy trawl gears in many shallow inshore parts.
Table 2.3.1 Length of shelf and the area of the shelf used in the analysis in the report.
|
|
SHELF |
0-20 |
20-50 |
50-70 |
70-100 |
0-100 |
100-150 |
150-200 |
100-200 |
200-500 |
TOTAL |
|
nm |
nm2 |
nm2 |
nm2 |
nm2 |
tot nm2 |
nm2 |
nm2 |
tot nm2 |
nm2 |
nm2 |
|
|
SURINAME |
185 |
3300 |
7750 |
2100 |
2300 |
15450 |
700 |
350 |
1050 |
2600 |
19100 |
|
GUYANA |
230 |
4200 |
6050 |
1750 |
1950 |
13950 |
800 |
800 |
1600 |
1500 |
17050 |
|
VENEZUELA ORINOCO |
90 |
2200 |
800 |
1500 |
850 |
5350 |
250 |
100 |
350 |
400 |
6100 |
|
TRINIDAD |
210 |
350 |
1050 |
900 |
1550 |
3850 |
900 |
850 |
1750 |
600* |
6200 |
|
JOINT AREA |
60 |
800 |
450 |
|
|
1250 |
|
|
|
|
1250 |
|
VENEZUELA ORIENTE |
240 |
850 |
4000 |
1750 |
3650 |
10250 |
900 |
900 |
1800 |
5400 |
17450 |
|
VENEZUELA WEST |
240 |
1100 |
900 |
|
1100 |
3100 |
1200 |
1200 |
2400 |
5400 |
10900 |
|
GULF OF VENEZUELA |
70 |
2100 |
1900 |
1350 |
500 |
5850 |
|
500 |
500 |
600 |
6950 |
|
COLOMBIA EAST |
490 |
550 |
2300 |
6750 |
1400 |
|
1400 |
3300 |
11450 |
||
|
COLOMBIA WEST |
490 |
2100 |
1800 |
6750 |
1400 |
|
1400 |
3300 |
11450 |
||
* Trinidad east coast only.The broad shelf off Suriname narrows west of Georgetown in Guyana and the gently sloping shelf is broken by a shallower bank some 20 nm from shore. This topographical feature seems to affect the fish distribution in the area.
Off the Orinoco River the soft mud can also extend onto the mid shelf. In the Colombus Channel, the bottom was found to be smooth and even, but soft in the southern parts.
Large parts of the bottom on the shelf to the east of Trinidad is uneven and rough. The Manzanilla Bank seemed to form a boundary with some areas of smooth bottom to the south. In deep water, 300-800 m the bottom was generally smooth, but with some rough and sharp stretches identifiable through echo sounder recordings.
The weather and current conditions on the Guyana shelf were generally favourable, but in the January-February survey a combination of strong currents and force 6-7 winds caused problems for trawling near the edge and on the slope.
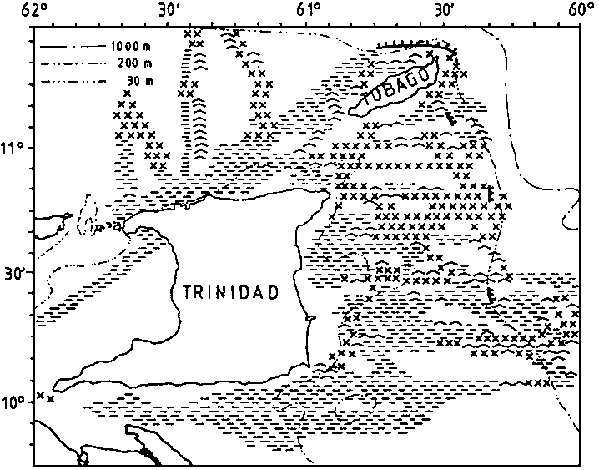
The topography around the Trinidad north coast and Tobago allows water from the Guiana current system to enter the Carribbean both in the shallower gap between Trinidad and Tobago and in the deeper parts north of this island. Westwards the shelf widens from about 40 nm at the north coast of Trinidad to 60 nm off Venezuela Oriente. The bottom depth increases fairly evenly out to the shelf edge at about 100 m of depth, but the character of the bottom is uneven and fairly rough over wide parts, especially offshore westwards to the Testigos Islands.
The entrance to the Gulf of Paria, the Dragon's Mouths may represent a feature of ecological importance because of its bottom topography and the effects of the entrance on the hydrographic environment
The bottom conditions round the Testigos and Margarita Islands were mostly even and smooth but with the slope outside the edge in part too steep for trawling. Not all parts of the narrow shelf past the Tortuga Island could be worked with trawl.
The well known Cariaco Trench has a threshold of about 200 m and anaerobic conditions in the deeper parts.
To the west of La Guaira the shelf becomes quite narrow, but with minor expansions in Golfo Triste and west of Istmo de Medanos. Except in the outer parts the Gulf of Venezuela has smooth even bottom.
Working conditions on the Venezuelan shelf were generally favourable. A prevailing easterly force 5-6 during surveys 1 and 2 did not affect the work in any significant way.
The Colombian shelf narrows westwards from the Guajira Peninsula and semi-gulfs and gulfs 30-40 nm wide shifts with parts of coast where the shelf edge is located only a few miles from the shore. The bottom over the shelf proper is even and smooth over most parts, but rough, uneven and very steep bottom was often encountered in the slope outside the edge of the slope. Strong easterly winds prevailed during surveys 1 and 2 and combined with high currents this made working conditions difficult near the shelf edge.
Figure 2.3.1. The bottom conditions as recorded by the acoustic system. (a)
Figure 2.3.1. The bottom conditions as recorded by the acoustic system. (b)
Figure 2.3.1. The bottom conditions as recorded by the acoustic system. (c)
Figure 2.3.1. The bottom conditions as recorded by the acoustic system. (d)
Figure 2.3.1. The bottom conditions as recorded by the acoustic system. (e)
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}